From the experiments performed by Takeuchi et al (1999) it was found that two genes of the T-Box family, Tbx4 and Tbx5, are important factors in determining the identity of limb buds in chicks. It is believed that the differential expression of Tbx4 and Tbx5 in leg and wing buds, respectively, are the determining factors where expression of Tbx4 tells a limb bud to form toes, scales, and the typical leg-skeletal pattern. On the other hand, the expression of the Tbx5 gene is prospective limbs influences the formation of wing-characteristic digits, feathers, and skeletal pattern.
However, as Takeuchi et al (1999), Logan et al (1998), Gibson-Brown et al (1998), and Rodriguez-Esteban et al (1999) show, there is a complex series of signals leading up to, and continuing after, the expression of the T-Box genes.
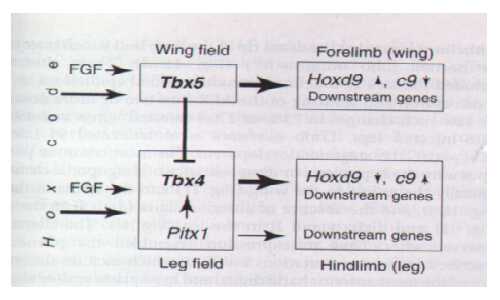
Figure 1. Genetic interactions involved in the expresion of the T-Box genes in prospective limb fields. The top pathway depicts the series of events taking place in the forelimb leading to wing development, whereas the bottom pathway represents the events leading to leg development in the hindlimb, (Takeuchi et al 1999).
In contrast, Tbx5 was confined to the forelimb and is detected by stage 14 adjacent to somite 10-23. This pattern of expression is shifted to somites 11-24 by stage 15.
Experiments performed by Takeuchi et al (1999) show that when RCASBP-Tbx5-EGFP or RCASBP-Tbx4-EGFP (see methods)are electroporated into prospective limb fields, misexpression caused dramatic morphological changes. As it was observed, when Tbx5 was ectopically expressed in leg fields caused elongation of the fibula including articulation with the metatarsal bones whereas the fibula is normally short and articulation does not occur. In addition, three digits were formed, versus four in normal legs, and partial scale to feather conversion. Thus it is clear that ectopic expression of Tbx5 induces partial wing-like morphology where a leg should normally form.
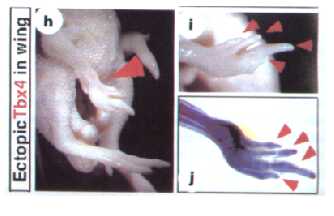
Figure 2. The effects of ectopically expressed Tbx5 in limb development. The red arrow in box D shows complete scale-to-feather conversion, whereas box E shows partial scale-to-feather conversion. Box F shows elongation and articulation of the fibula (red arrow) contrasted with a normal leg (box G), (Takeuchi et al 1999).
Similarly when Tbx4 was introduced to presumptive wing-fields, a partial leg-pattern was observed with four toe-like structures, complete with claws and feather-to-scale conversion. In addition, some skeletal deformations were observed, including thinning of the ulna.
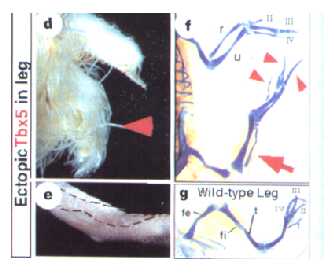
Figure 3. The effects of ectopically expressed Tbx4 in limb development. The red arrow in box H shows complete feather-to-scale conversion. Box I shows four separate digits with claws, while box J shows the thin ulna along with the four complete digits, (Takeuchi et al 1999).
As described in the interactions page, the ectopic misexpression of one T-Box gene utimately interferes with the expression of the other. Consequently, the observations of endogenous Tbx4 transcripts were down regulated as Tbx5 was ectopically introduced. On the other hand, Tbx4 cannot suppress endogenous Tbx5 expression thus demonstrating a hierarchy of sorts.
This phenomenon can be explained through the interpretations of experiments performed by Takeuchi et al (1999), Rodriguez-Esteban et al (1999), and Johnson and Tabin (1997) involving the fibroblast growth factor-8 (FGF-8) induced extra limb system. When FGF-8 is expressed in prospective limb buds close to somite 21, extra wings developed, whereas ectopic FGF-8 close to somite 25 induced extra legs to develop. However, when FGF expressing cells were mixed with TBx4 producing cells and implanted at somite 21, endogenous transcripts were down regulated and Tbx4 transcripts were expressed throughout the limb bud, influencing structures to form as a leg would. As described later, expression of the Hox gene, Hoxc11, normally expressed only in legs, were normally observed.
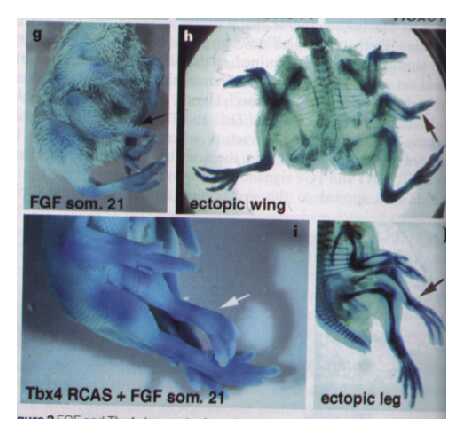
Figure 4. The effects of ectopic FGF transplants on limb development. Ectopic expression of FGF alone give rise to extra wings (box G and H), whereas box I and J show the development of an extra leg when FGF is coupled with Tbx4 expression, (Rodriguez-Esteban et al 1999).
Thus, a hierarchy of expression is theorized with Tbx4 acting as a major determinant of leg identity where leg development is a more likely result when Tbx4 is expressed early in development. In other words, Tbx4 can induce leg-like morphology, even in wing-destined limb fields, when expressed at the initial stages of limb development, whereas the influence in later stages is not as great and limb conversion is less likely.
The T-Box genes are involved with a number of gene groups, including the Hox code and Pitx1. When Tbx5 is misexpressed in limb fields, wing specific Hoxd9 was clearly induced, coupled with repression of endogenous leg-specific Hoxc9. Tbx4 has a contrasting effect where Hoxc9 was misexpressed in wings, with the normal Hoxd9 gene being down regulated.
The gene Pitx1 seems to have some interactive role with Tbx4 in achieving leg identity as the expression of Pitx1 was not affected in either misexpressed Tbx5 legs or Tbx4 wings. Pitx1 is seen normally in legs by stage 11, preceding the expression of the T-box genes (stage 14 for both). The initial expression domain of Pitx1 is fairly broad, not being confined to the prospective leg fields, as is Tbx4, implyinga role in inducing Tbx4. Takuchi et al (1999) tested this hypothesis by implanting FGF producing cells at somite 23 and observing the sequential order of expression of genes. It turns out that Pitx1 is expressed and observed 6 hours after implantation, whereas the T-Box genes are observed 12 hours later.
However, it was noticed that the Tbx4 being expressed was done so adjacent to the area of FGF producing cells, leaving a gap in the expression domains. This observation implies that the Pitx1 gene acts to regulate the TBx4 indirectly.
With regards to early limb development and the expression of the T-Box genes, Gibson-Brown et al (1998) observed the involvent of such with respect to the AER and ZPA. When leg mesenchyme cells were grafted beneath the AER of the wing bud, the misexpression of Tbx4 was observed. Similarly, when Tbx5 producing wing cells were grafted beneath the leg AER, Tbx5 gene transcripts were clearly observed.
Interestingly, when the AER was removed in developing wing and leg buds, the expression of Tbx5 and Tbx4, respectivley, were maintained. Thus, the maintenance of the T-Box genes, once established, are not dependent of the continuation of the AER/ZPA derived signals.
It has been discovered that mutations in the Tbx5 gene are responsible for causing human diseases such as the Holt-Oram Syndrome (Gibson-Brown et al 1998 and Logan et al 1998), affecting the development of the fore-limb. The Holt-Oram Syndrome is characterized by a reduction in the radius and loss of the thumb. As it is, there are no phenotypic diseases or deformations known yet, caused by mutations involving the Tbx4 gene.
In conclusion, the role of the Tbx4 and Tbx5 gene is clearly that of an important one in the identification process of limb buds. It shows that the T-Box genes regulate the expression of some downstream targets that influence the formation of structures that are directly related to the specific T-Box gene being expressed. In other words, Tbx5 and Tbx4 induce downstream factors to follow a pathway that makes wing-like structures in Tbx5 expressing limbs versus leg-like structures in Tbx4 expressing limbs.
Go to
The methods used The Hox genes Glossary of Terms Gene Interactions References
Go to the
Top
