
The sequence of events that begins with a simple collection of cells in an early embryo and ends with a fully formed leg or wing is a complicated one involving multiple signalling cascades, including numurous gene families such as: the Hox gene family, FGF's, the T-Box genes, and genes involved in AER and ZPA formation.
As stated before (see early limb development), the AER is formed from the convergence of dorsal ectoderm, including the somites and lateral plate, which solely expresses En-1, and the ventral ectoderm containing the r-Fng gene. It turns out that the interplay between these two genes determines the exact position where the AER forms, that is, the boundary line between the r-Fng expressing cells and the non-r-Fng expressing cells. The mode by which this is achieved is similar to the way the main body pattern is laid out in that the En-1 represses the expression of r-Fng, thus forming a sharp boundary between the dorsal (r-Fng cells) and ventral (non-r-Fng cells) ectoderm (Johnson and Tabin 1997, and Logan et al 1998).
In addition, the En-1 gene is also involved in functioning as a ventral regulator influencing dorsal/ventral patterning. It seems (Johnson and Tabin 1997) that En-1 act to repress the expression of the dorsally expressed Wnt-7a ventrally such that, in turn, Wnt-7a can confine the expression of the LIM-homeodomain protein Lmx-1 to the dorsal mesenchyme. Then, by some unknown mode of action, Lmx-1 instructs dorsal patterning in limb bud mesenchyme.
Figure 1. Genetic interactions involved in the formation of the AER on the dorsal/ventral ridge (left pathway) and in determination of the dorsal pattern (right pathway), (Johnson and Tabin 1997).
In addition, the Wnt-1 gene along with two other gene groups, BMP and FGF, are intrinsically linked to the T-Box genes (Rogriguez-Esteban et al 1999) such that all three gene transcripts were down regulated when either T-Box gene was ectopically misexpressed. Conversly, when, for example, the gene BMP was artificially expressed in leg bud fields, the appearance of Tbx5 was observed some time later; when the expression of BMP was blocked, the appearance of Tbx5 transcripts were diminished.
As mentioned before (see the T-Box genes), the group of FGF factors, most noteably FGF-8 and FGF-10, are closely involved with the T-Box genes in influencing limb development. As observed by Johnson and Tabin (1997) and Logan et al (1998), the gene FGF-8 was deemed an excellent 'candidate' for playing a part in limb bud initiation as it is localized in intermediate mesoderm at the fore and hind-limb levels prior to limb bud formation. In the same aspect, it was shown that FGF-8 when introduced ectopically can initiate an entire new program of development.
However, it was later found by Gibson-Brown (1998) that FGF-8 merely acts in a relay, initiating a second set of signals starting with FGF-10.
It was shown by Gibson-Brown (1998) that the Hox genes and the T-Box genes interact to influence skeletal patterning. Through observations of gene transcript expression, a pattern was observed such that:
To summarize, regions that expressed Tbx5 develop wing digits and feathers whereas regions which express Tbx4 develop toes and scales. It is thought that this patterning is controlled by members of the Hox code being expressed at different times during development.
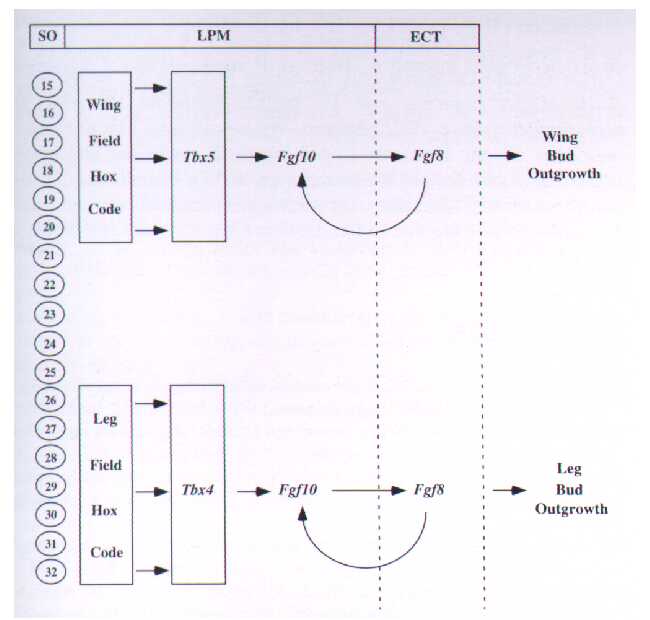
Figure 2. Flow chart summarizing the interactions between genes involving in limb identification and outgrowth (Gibson-Brown et al 1998). The vertical bar on the left side depicts the somite level (SO), the intermediate section depicts the interactions of signals in the lateral plate mesoderm (LPM), and the right hand side refers to actions in the ectoderm (ECT).
The individual T-Box genes themselves interact with one another, such that cross talk between Tbx5 and Tbx4 allow for autoregulation within the promoter of the specific T-Box genes, such that Tbx5 can supress Tbx4 in wing fields and vice versa. In addition, Rodriguez-Esteban et al (1999) hypothesized that since the T-Box gene products bind the DNA as homeodimers, the ectopic expression of another T-Box gene within the same limb field may cause the formation of heterodimers between the endogenous and ectopic proteins. This in turn could inactivate the endogenous proteins, preventing DNA bindng and activation of downstream targets.
Thus as it can be seen, the T-Box genes, Wnt, BMP, and members of the FGF's, Hox genes, and AER/ZPA systems form an intricate web of signalling patterns and feedback loops, most of which is still unknown.
Go to
The T-Box genes Discussions The Hox genes Glossary of Terms References
Go to the
Top
