The Theory of Island Biogeography
Island biogeography has been a subject of
considerable interest to biologists and geographers since the time of Darwin,
Wallace, and the less well-known Hooker. Hooker explored islands in the South
Atlantic and South Pacific. Darwin and Wallace are more important in our
current thinking, since these two were pioneers in the development of the
theory of evolution. However, much of the theory of island biogeography was
built on data which came from their studies of the Galapagos and East Indies
respectively. Islands have been studied as natural experiments ever since, with
varying levels of intensity. Oceanic islands are isolated and small enough to
reveal processes and results too complex to interpret in mainland areas.
Islands are unique. Since they are isolated,
evolutionary processes work at different rates - there is little or no gene
flow to dilute the effects of selection and mutation. Endemism is rampant.
However, both in theory and practice, that same isolation makes islands more
vulnerable to habitat change and extinction. Introduction of a single predator
or herbivore can have dramatic impact on the local community, as we have just
seen with biota of the Hawaiian islands.
Islands, as natural experiments, have not
been protected from damage and extinction through human activities. Since
islands are isolated, and in many cases the species found on them are endemic,
extinction has been particularly common on islands. 93% of the bird species
whose extinction has been recorded since 1600 have been island species.
Historical records suggest a mean extinction rate through the Pleistocene of
approximately 1 species in 83.3 years. In 1980, that rate was one species every
3.6 years. Extrapolating the curve, an extinction rate of one island species
per year can be predicted for the immediate future.
One of the reasons islands are important in
the more general structure of ecology, biogeography, and conservation biology
is that islands, as at least relatively isolated areas, are excellent natural
laboratories to study the relationship between area and species diversity. When
we fully understand the relationship, it will be applicable to fragments of
habitat that human activities protect. We all know those sanctuaries are
important, but we need to know what and how much we can protect in them.
Species-Area Relationships
One of the immediately obvious
characteristics of islands is the number of species resident there, a number
much lower and therefore more countable than the diversity on a continental
mainland. Such counts can reveal interesting relationships. For example, Great
Britain has 44 species of mammals, yet Ireland, only approximately 20 miles
further removed from mainland Europe into the Atlantic, has only 22 species. Is
20 miles a sufficient distance to increase isolation and decrease mammalian
immigration by half? If so, then flying mammals (bats) should show similar
numbers of species on both islands, since immigration and isolation would be
significantly less severe to a bat species. The number of bats is not similar.
Only 7 of the 13 bat species resident in Great Britain breed in Ireland. What
factor accounts for the difference? The single factor which provides the best
explanation is island area (though it is not the only contributing factor).
There is a classic curve originally drawn by Darlington, and reprinted in every
biogeography chapter since then, which depicts the relationship between area
and the number of reptilian species in the West Indies.
Figure
- A curve relating island area to
reptilian diversity in the West Indies.
As a log-log plot it is not a 'curve', but a
straight line. From it a 'rule-of-thumb' can be formulated which says that
increasing the area of an island by a factor of 10 (or, more correctly,
counting the number of species on a nearby island 10 times larger) would
approximately double the number of reptile species present. That relationship
is called a power function. For islands it is written as:
S = C Az
where:
S is the number of species present,
C is a constant which varies with the taxonomic group
under study (taxa which consist of good dispersers (these species also
typically have rapid population growth) will logically accumulate more species
on an isolated island, all else being equal),
A is the area of the island, and the exponent z has
been shown to be fairly constant for most island situations.
Geographic variation in C
has been observed and 'loosely' reflects the isolation of island groups
typically studied due, for example, to the presence or absence of major air or
water circulation pathways nearby (islands located along the Gulf Stream would
be more likely to accumulate species than those located in the doldrums of the
Sargasso Sea) and also to effects of gross climatic difference (i.e. C is
higher in the tropics than for islands at high arctic latitudes). C can also be
regarded as a scaling factor. In terms of the graph of a species-area
relationship, z determines the 'shape' of the curve, which is raised or lowered
as a whole by the value of C. The meaning of z, in an all out treatment, is
related to the distribution of abundances of species, i.e. to the number of
species expected if the total number of individuals increases (as it would on a
larger island) and those species follow a Preston log-normal distribution of
abundance (see May 1975 for a full treatment). However, there is a simpler
level of meaning for the z, revealed by manipulating
the equation. The graph of the basic equation would show
an exponential rise in the number of species as area increased. If we take the
logarithms of both sides of the equation, we get something that should look
easier to work with:
log S = log C + zlogA
If we conveniently forget
that the equation has logs as its terms, then you can see that this is the
equation for a line, with the basic formula:
Y = b + mx
So
that if we plot the log of the number of species against the log of island area,
we get a straight line, and the slope of the line is the coefficient z. The z,
establishes the characteristic relationship between the number of species and
area for all taxa. It is a general characteristic, and fittingly, it is related
to the most basic hypotheses about the way species are put together in
communities. There is variation in z, but it reflects, in theory at least,
general patterns of isolation involved in the studies. Study of taxa in
mainland 'islands' less isolated from their surroundings will give a lower z
value (though much higher total species numbers) than would parallel studies of
the same taxa on oceanic islands. It is argued this results from species that
could not live indefinitely within the 'island' being observed as transients in
mainland habitat 'islands', though they would never be observed on oceanic
islands. Is z really as independent of autecological
characteristics of species and effects of real world disturbance as this view
might lead you to think? The answer, at least from modeling studies to be
considered below, is a clear no. The value of z seems to vary with the growth
characteristics of species and with the frequency and intensity of disturbance
(Villa et al. 1992).
Species-area curves have been generated for a
large variety of places and taxa, and the range of z values is remarkably
small. The data in the table below fit fairly closely with what would be
predicted based upon the theory of species abundance distributions. May (1975) gave an extended treatment of the mathematical relationships
among observed and theoretical abundance and diversity patterns. Here the
treatment will be limited to a more compact form (derived from a chapter on
patterns in multi-species communities in May (1976)). The most commonly observed
distribution of the relative abundance of species is a log normal. This means
that a plot of the logs of species abundance against the number of species
(i.e. a histogram) follows a bell shape. Sampled distributions usually have
what is called a 'veil line'.
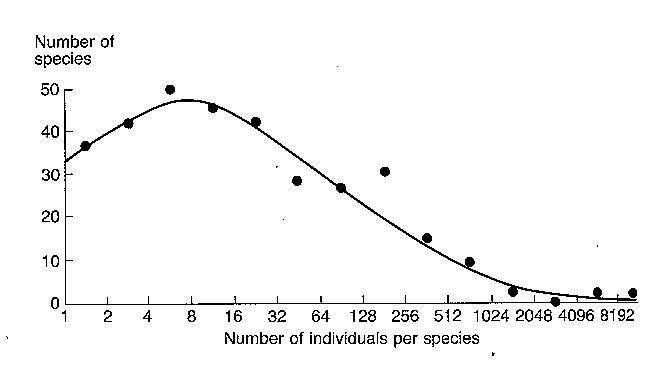
Figure - Lognormal abundance distribution
for moth samples.
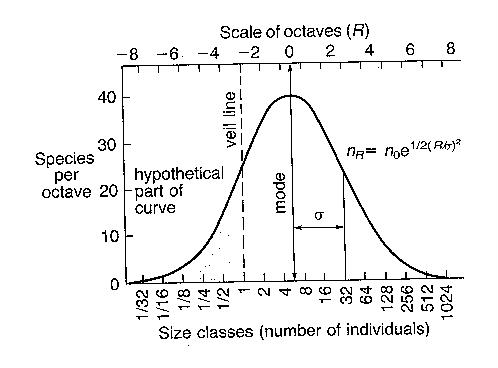
Figure
- The theoretical Preston log-normal species abundance distribution
The curves indicate the presence of a few
common species (the right hand end of the curve) and a larger number of species
of intermediate abundance. We don't usually see the left hand end of the curve
(the very rare species) because we rarely sample enough individuals to capture
even one. A larger total sample moves the veil line to the left, taking in more
of the total curve. Preston used other data, a log-normal species abundance
distribution for birds in Maryland, to develop the log normal.
Theoretically, the Preston
log-normal the z value should be 0.263.
May says that the problem is that z doesn't really tell us much, since
within the constraints as S varies from 20 to 10,000 species, the z value
changes only from 0.29 to 0.13. It really turns out that z is largely a
mathematical property of the log-normal, and doesn't tell us much about
communities. If they are log-normal, then z will be at least close to .263. Nevertheless,
there is still some interest in patterns in z. To indicate that briefly, here's
a short table of some species-area relationships:
Group Location value
_____________________________________________________________
beetles West Indies 0.34
reptiles and West Indies 0.30
amphibians
birds North America-Great 0.17
Basin
Birds West
Indies 0.24
land vertebrates Lake Michigan 0.24
Islands
ants Melanesia 0.30
birds East Indies 0.28
land plants Galapagos 0.33
dipterans Cincinnati parks 0.24
mammals North America 0.1-0.2
The values of z for mainland areas are
clearly lower, e.g. both diptera in parklands and
mammals over North America generally, than the values for isolated islands. As mentioned
above, the explanation for these lower values is the inclusion of transients in
species counts from small 'islands'. Consider a badger, with a home range of
200 square km, or a wolf with a home range of 400 square km, or even larger
areas for seasonal migrants like caribou or large predatory birds. These
animals could easily be included in counts from 'island' areas much smaller
than their home ranges, i.e. they could be present in a 'patch' of mainland
area considered as an 'island' transients. They could not survive in such small
areas if isolated by significant barriers, but sampling larger areas more
comparable to their home ranges would then not add them to species lists as if
new; they are already on the species list from being present as transients in
the smaller areas. Oceanic islands do not contain such transients. Thus we get
measurements of species numbers which are 'too high', include transients, in
small mainland sample areas, and too few additions as larger areas are sampled.
The slope of the graph relating the number of species to island area, i.e. the
z, is lowered. The theory underlying the log normal and island biogeography
implicitly assumes that all species counted could be resident permanently
within the 'island' area, this may not be the case
with some mainland species. Compare the slopes for bird species numbers on
islands in the East and West Indies, both >0.2, with that for boreal forest
birds in the Great Basin of western North America (0.16). All three values are
for habitat islands, but on mainland the slope is lower, and the likely reason
is the presence of some 'transient' birds in species counts within small
habitat islands in the Great Basin. That may also explain the low value for
mammals, but what about the low value for Cincinnati flies? Are they likely to
be transients? The explanation here more likely lies in the assumption that
islands are truly isolated; immigration onto an island requires a jump dispersal, i.e. the intervening habitat
is assumed to be inhospitable. The same sampling artifacts that led to low
values for mammals do also apply here. Models assume island populations are
numerically self-sustaining, and have increasing probabilities of extinction on
the island as they become rarer. Other green areas, garbage, etc. may sustain
fly populations (and even permit reproduction!) in habitats between park
'islands' in the urban area, permitting high rates of re-immigration, so that
populations which might not otherwise be sustained in a park remain. That 'rescue'
is much less likely for oceanic island populations.
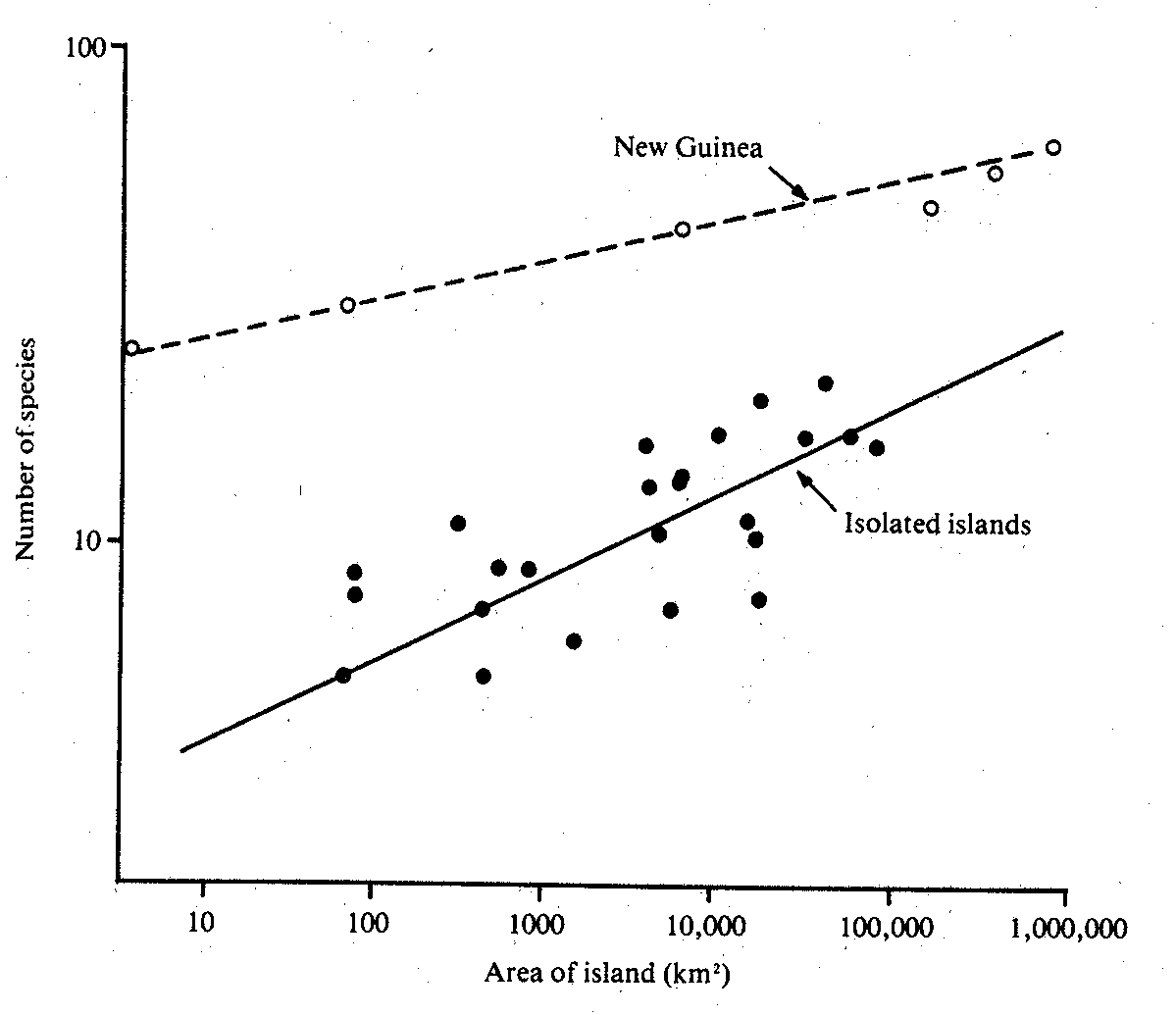
Figure
- Species-area curves for ants on New Guinea (relatively, a mainland) and the
isolated islands nearby. The island curve is steeper (a higher z) than the New
Guinea curve, as explained above.
Is the relationship between
species and area linear?
There are theoretical
reasons to expect a z of 0.263. However, in doing so we are accepting that
species-area curves are linear. There are some points to consider:
Given apparent differences in what you might
expect, is the linear equation predicting z logical? There is at least one
study that questions the assumption of linearity (Crawley and Harral, 2001). They sampled Berkshire and the East Berks in
England in nested, contiguous quadrats, at 100 and
25km2, also 456 km2 contiguous quadrats
and replicates (not contiguous) from 0.01 m2 to 110 ha in Silwood Park. Initial results, averaging the values
obtained, don't differ significantly from the theory. They found z=0.302 for samples from the Silwood research estate, and z= 0.267 for samples over the
larger area of Berkshire and all of Great Britain.
An important point about effects of area on
species diversity may have slid by here. The biological question is why does
area affect species numbers? There are two schools of thought:
1) The
Preston canonical log normal distribution can be used to suggest that area
determines the total population size of the collection of species living there.
Area is the direct determinant of diversity, since the multiplicity of factors
which determine relative abundance and species diversity are prescribed, and
independent of the specific island of area being studied.
2) The
alternative school suggests that area is of only indirect importance. Area fits
because area is a good indicator of the amount of habitat diversity present on
an island. It is really the 'number of niches' that determines the number of
species, but there is no established method for counting, or even estimating
the number of niches in an environment. Instead, physical variables are usually
measured. The assumption is made that the two measures should be highly
correlated.
Let’s look at some of the basic data used to
verify the basic relationship between species numbers and island areas. One of
the frequent approaches is the use of multiple regression models to determine
what factors account for differences in species diversity on islands. Jared
Diamond (1973; Diamond and Mayr 1976) used this method to study the
distribution of bird species on New Guinea and its satellite islands. The data
were first fit to the basic equation; for these bird data the equation then
reads:
S
= 15.1 A22
for the satellite
islands near New Guinea, and this relationship accounted for 81% of the
variation in the number of bird species among islands studied. When Diamond
instead measured the numbers of species in montane habitat islands on New
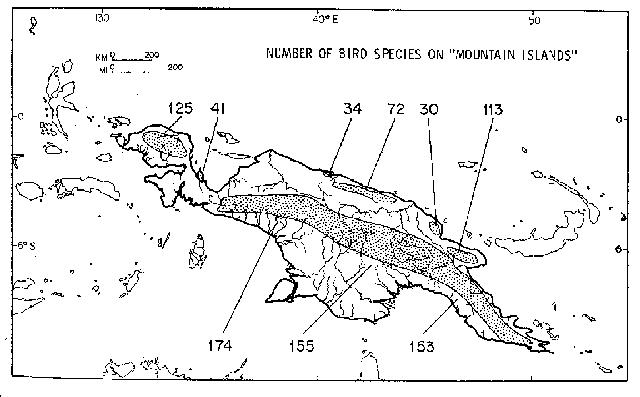
Guinea proper, only the constant changed, to 12.3. Figure 4 shows the numbers
of species and the locations of montane habitat islands. The central 'line' of
New Guinea is a mountain backbone, and forms the largest single island. Diamond
discerned that a part of the effect of increasing area was due to an increase
in the maximum elevation observed on larger (even habitat) islands, and the concommitant differences in the number of habitats
resulting from a greater range of elevations, which cause differences in
climate.
Figure
- A map of the montane areas of New
Guinea and the diversity of montane birds in those areas. Variation in species
numbers is correlated with variation in altitude.
In addition to the number of species
accounted for by area alone, each 1000m of elevation 'caused' an increase of
2.7% in bird species diversity on New Guinea satellite islands (and 8.9% in New
Guinea habitat islands) on average. After the entry of this term, the
regression equation reads:
S
= 15.1 (1 + .027L/1000) A22
Diamond realized that the number of species
on islands also reflected the degree to which the islands were isolated by
distance from source populations. The more distant from the New Guinea source
of species, the smaller the number of species when islands similar in size and
elevation were compared. The decrease is approximately exponential with
distance, and the number decreases by half for each 2600 Km on average. Thus,
Pitcairn Island (the place where the Bounty's mutineers ended up) is about 8000
Km from New Guinea, and has about 1/8 or 12.5% of the bird species numbers on a
similar area of New Guinea. When this factor in incorporated into the multiple
regression equation, the result is:
S
= 15.1 (1 + .027L/1000) (e-D/3750)
A22
The following figures show you how some of
the data look, and why the distance relationship is exponential. Figure 5 shows
the basic species-area relationship for the satellite islands around New
Guinea.
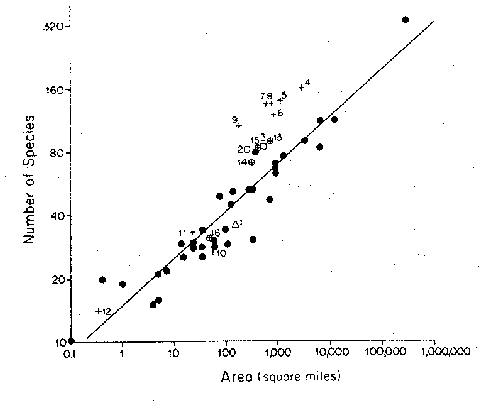
Figure
- Species-area relationship for birds of
oceanic islands around New Guinea. Differences in isolation are evident both in
absolute numbers of species and in different slopes for islands in different
distance ranges.
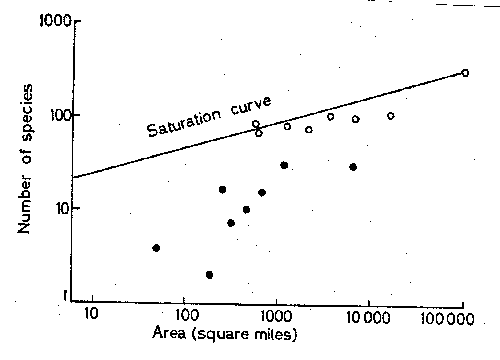
Figure
compares some of the data for satellite
islands to the curve for areas of the New Guinea mainland (called a saturation
curve). The slope is clearly steeper for the islands than for mainland areas of
similar habitats. The open circles are points for islands relatively near New
Guinea, the filled circles for more distant islands. More distant islands have
fewer species for the same area.
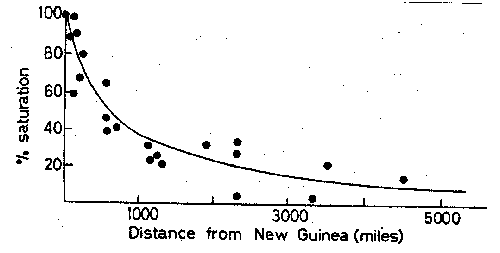
Figure
looks at that relationship. Each island diversity is assessed as the fraction of the
number of species expected in a mainland habitat of the same area. This %
saturation is plotted against distance from New Guinea.
Other
data sets have been analyzed using the same basic approach, but one can significantly
add to our understanding of the biology underlying island diversity patterns.
In a study of bird species diversity on the Channel Islands off Santa Barbara,
California, Power (1972) worked backwards from the physical variables to the
biological variables which correlated most closely with bird species diversity,
using a method called path coefficient analysis. The method begins with a
multiple regression analysis paralleling the study of New Guinea satellite
islands. Power first found the factor which explained the largest portion of
the total variation in bird species diversity, entered it into the model
equation, then determined the factor which explained
the largest portion of the variation which remained after applying the model
equation. That factor enters second, and forms part of a new model equation.
Subsequent factors are entered in order of importance, using the variation
about the model equation fitted using factors already incorporated. In order of
importance, Power found the number of plant species on islands was the best
single predictor variable, accounting for 67% of the total variation in bird
species diversity. Island area, on the other hand, accounted for only 33% of
variation if considered alone. It seems logical that habitat heterogeneity (or
diversity) for bird species would be measured by the biotic diversity of their
residences, food sources, or residences of their food resources, i.e. the
diversity of the plant community.
Having
used plant species diversity as the first factor entered stepwise into the
model, what was the next most important factor? Not area! Once effects on plant
species diversity are removed, area doesn't even account for a significant
fraction of remaining variation, let alone be the most important factor in
residual variation. Only one factor is significant in explaining residual
variation of a model including plant species diversity. That factor is
isolation (distance), which accounts for 14% of total variance when entered as
the second factor in the stepwise model equation. The remaining 19% of variance
is unexplained variation, called error variance; no other factor accounts for a
significant portion of variation out of this residual. This is a solid
indication that measures of habitat heterogeneity (diversity) are the critical
predictors of island species diversity.
Power,
however, went further. Since plant species diversity, used as a predictor
variable (or factor) is itself a biological variable, he asked what, in turn,
explains plant species diversity. Using the same stepwise regression method,
Power found 3 factors were significant, and they were 3 factors that correspond
closely to those which Diamond found significant in explaining bird species
diversity in the New Guinea satellite islands: 1) area accounts for the largest
proportion of plant species diversity, 68%; 2) latitude accounts for 15% of the
variance. Latitude effects are detectable only after effects of area have been
removed, especially since the latitudes of the Channel Islands don't differ by
much. This factor apparently measures position of islands relative to
North-South air and water currents off the California coast, rather than a
direct impact of latitude itself. 3) Isolation, or distance from the nearest
source pool, with the islands all at similar distances from mainland, accounted
for only 3%, but was significant. The remaining 14% of variation was
unexplained, i.e. error variance. Taking this analysis at face value, we can
see the origin of path coefficients. It suggests that the underlying factors
which explain bird species diversity on the California Channel Islands are
abiotic, the same factors which Diamond found in his studies, particularly area
and factors important in assessing isolation, but that the proximal factors important
to the birds are biological as well. A statistical path then takes the
following form:
area
----> plant species diversity
latitude island area
----> bird species
distance isolation diversity
At this point it should be clear that a
major group of biogeographers believe that island area is an excellent, though
indirect, indicator of island species diversity.
While we may be accustomed to thinking of
islands strictly in 'geological' terms, it is clear that islands take many
forms, including lakes, forest patches in agricultural lands, or even zebra mussels colonies. The key aspect of islands is that they are
favourable habitat surrounded by inhospitable
habitat. Looking at how zebra mussel colonies on soft sediments in Lake Erie
can be 'islands', Bially and MacIsaac (2000) looked at invertebrate species
diversity in relation to island area. A very clear image emerged. Small islands
(10 cm2) host only about 8 species, on average, while midsize islands
(100 cm2) supported ~13, and large islands (70,000 cm2)
supported about 20 taxa. The invertebrates utilize gaps between mussel shells
as habitat, and mussel feces and pseudofeces as food.
The Basic Model of Island Biogeography
The model is one of a dynamic equilibrium
between immigration of new species onto islands and the extinction of species
previously established. There are 2 things to note immediately: 1) this is a
dynamic equilibrium, not a static one. Species continue to immigrate over an indefinite
period, not all are successful in becoming established on the island. Some that
have been resident on the island go extinct. The model predicts only the
equilibrium number of species, will remain 'fixed'. The species list for the
island changes; those changes are called turnover. 2) The model only explicitly
applies to the non-interactive phase of island history. Initially, at least, we
will consider only events and dynamics over an ecological time scale, and one
which assumes ecological interactions on the island occur as a result of random
filling of niches, without adaptations to the presence of interacting species
developing there. Evolution is clearly excluded.
The
variables used in the basic model are Is,
the immigration rate, which is clearly indicated by the subscript to be species
specific, i.e. to be dependent on the number of species already present on the
island. Here we're not counting noses, but rather the rate at which new species
(those not already present on the island) immigrate. Phrased explicitly, it is
the number of species immigrating per unit time onto an island already occupied
by S species. Also Es, the extinction rate, measured in species lost
per unit time from an island occupied by S species. Finally, we need to know the
size of the pool of species in the source area available to colonize the
island.
The immigration rate Is must certainly
decrease monotonically (on average) as the number of species on the island
increases, since as S increases there are fewer and fewer species remaining to
immigrate from the pool P of potential immigrants at the source. If all species
were equally likely to immigrate successfully (i.e. had equal dispersal
capabilities), but actual immigrations were chance events, then the relationship
between Is and S would be linear, the
probability of a new species immigrating would be directly proportional to the
number of species left to arrive. There are, however, considerable differences
in the dispersal abilities of species in source areas. Those with the highest
dispersal capacities are likely to colonize an island rapidly (have a higher
immigration rate), and later, on average, those with lower dispersal capacities
will follow. They will not only immigrate later, but the rate at which they immigrate
will be lower because they have lower dispersal capacities. The rate at which
species accumulate on islands is, therefore, initially rapid and then slower.
Also, among those species with lower dispersal capacities the successful
immigration of any one species should have less effect on the immigration rates
of species remaining in the source pool (we have not removed a likely immigrant
from the pool) than would the earlier immigration of a highly dispersible
species. Therefore, this part of the curve should be 'flatter'; the rate of
immigration should be little affected by the arrival of one of these poor
dispersers. The result is an observed immigration rate curve which is concave.
The actual (or theoretical) curve for any island is dependent on its isolation.
For any source pool, the observed rate, while similar in shape, will be lower
for more distant islands than for closer ones. Immigration rates are graphed
from the left hand edge of figure 1, declining from the y axis with an
increasing number of species already present.
Figure
- The basic graphical model of equilibrium in the MacArthur-Wilson model.
Figure from Brown and Gibson -Biogeography.
The extinction rate Es should be,
from parallel reasoning, a monotonically increasing function of S. If area, for
example, acts only through its effects on population sizes, and extinctions are
the chance result of small population sizes and demographic stochasticity,
then as the number of species increases, the number of species with small
populations and subject to chance extinctions increases in proportion, i.e. the
relationship would be linear. However, if we consider a more realistic
biological scenario, then as the number of species increases, depressant
interactions within and between species (competition, predation, parasitism) are more likely to occur, and extinctions are
more likely as a result. Remember that these are not species that have evolved
adaptations to interactions. Effects are direct and unmoderated.
Since any extinctions resulting from interaction are in addition to those
resulting from demographic stochasticity, the more
realistic shape for the extinction curve is concave upwards. The extinction
rate begins at 0 when there are no species on the island, then
increases as species accumulate. At least for purposes of simplicity in looking
at the basic implications of the model, the extinction curve can be thought of
as a mirror image of the immigration curve.
We
now have all the information to produce the basic graphical model. That model
predicts that there is some value of S, which is called Ŝ, for which
immigration rate and extinction rate are in balance; there is a dynamic
equilibrium. At that diversity on the island species are immigrating at a rate
equal to disappearances due to extinction. The result is constant change in the
species list on the island; that change in names occurs at a rate called x, the
turnover rate. The length of the species list, however, should remain constant.
This is a stable equilibrium since, should something happen, and the number of
species on the island be perturbed, the imbalance between immigration and
extinction rates at the new S would tend to return island diversity toward its
equilibrium value. Below Ŝ additional species accumulate; immigration rate
is larger than extinction rate. Above Ŝ the reverse is true, extinctions
exceed immigrations and the number of species declines to Ŝ.
Tests of the Model
To
test the model, an important piece of evidence is a carefully designed
manipulative experiment studying the fauna which colonize 'islands'. One of
Wilson's students, Dan Simberloff, tested the model using islands which consist
of mangrove mangles in the Florida Keys. Simberloff's
Ph.D. thesis had consisted of measurements of the re-colonization of these
islands following 'defaunation' (he had encased
individual mangles in giant plastic bags, sprayed them with short acting, low
persistence insecticides, then followed the rates, numbers, and species which
immigrated onto them after exposure). Re-equilibration, i.e. reaching a stable
number of species, had occurred within 3 years of fumigation in his earlier
experiments. In a second series of studies (Simberloff 1976), the manipulations
were equally inventive. After the
islands had been censused, and an equilibrium number
of species determined for each island (a 'control' diversity), crews moved in
with chain saws, handsaws and hatchets, and each island was split into 2 or
more smaller parts, with water gaps of 1m between. To the insects, apparently
this 1m gap was sufficient to make crossing from one sub-island to another a jump dispersal. The smaller, sub-islands were then censused repeatedly over a time interval sufficient to
permit re-equilibration to find out how species numbers changed with island
area. Remember, the area censused had been part of a
previous island, and should contain all habitats (plant parts, vertical
structures) in the same proportions as before (i.e. the same habitat
heterogeneity, however measured). Alterations were only quantitative, in the
form of area reduction, no unique feature was removed.
The
results were clear-cut. Each island reduced in size re-equilibrated at a lower
insect diversity. Considering all the experimental islands in developing a
model for the pattern in reduction, the diversity change fit a log-log
relationship (i.e. a power function) between diversity and area. Thus, Simberloff's data fit the original species-area
relationship. Area was the key determinant. The process of re-equilibration,
however, involved extinction of species from islands supersaturated due to
their reduction in size. We have already encountered the underlying biological
cause of those extinctions: population sizes of 'marginal' species,
that is those whose populations were already small before reduction in
area, were decreased to the point where chance extinction due to demographic stochasticity became likely, and re-colonization unlikely.
Such extinctions are an important component of the equilibrium model of island
biogeography.
Figure
- Effect of island fragmentation on insect diversity in mangrove mangles. Simberloff (1976).
There
are few islands that have been studied over long enough periods to test the
hypothesis of equilibrium with turnover, i.e. the occurrence of a stable but
dynamic equilibrium. Among those few are the California Channel Islands. The
interpretation of these data is a source of continuing controversy. That's
important, because the crux of the equilibrium theory is proof (or
documentation) of insular turnover at equilibrium. A paper (Gilbert 1980) found
25 attempts to document turnover at equilibrium, and found few (basically just
mangrove island studies by Simberloff) acceptable without question. In Simberloff's original defaunation
studies, for example, one island supported 7 species of Hymenoptera prior to
fumigation and 8 after equilibrium had been re-established about one year
later. However, only two of these species were present both before and after
fumigation. This sort of experimental study is designed to allow for rapid
re-equilibration.
The
Channel Island studies represent an interesting attempt to deal with the
problems of scale (here time) when dealing with most real ecosystems.
Recognizing that there may be difficulties (the initial, historical survey of
species presences on the island used breeding records collected over many
years, rather than a single survey at one time), Diamond's studies of turnover
on the Channel islands are still regularly cited (Diamond 1969).
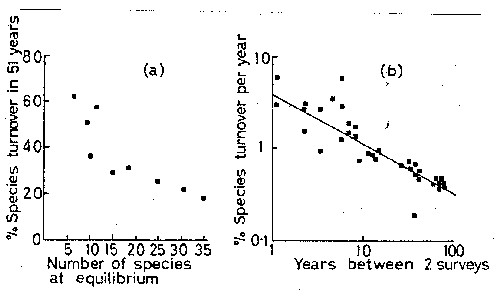
Initial
data reported collections and observations indicating the fauna of individual islands
in 1917. Diamond compared those species lists with a survey he did in 1968.
Over the 51 years between censuses the numbers of species on islands remained
almost perfectly constant, but turnover was as high as 62%, i.e. as much as 62%
of the original list had been replaced by new species. The islands had the
following characteristics:
Island 1917 1968 Extinctions Immigrations %turnover
Los Coronados 11 11 4 4 36
San Nicholas 11 11 6 6 50
San Clemente 28 24 9 5 25
Santa Catalina 30 34 6 10 24
Santa Barbara 10 6
7 3 62
San Miguel 11
15 4
8 46
Santa Cruz 36
37 6 7 17
Anacapa 15 14 5 4 31
These data seem initially to
fit the equilibrium theory quite well. Numbers remain almost constant while
turnover occurs in a significant number of species. However, the theory also
suggests, as you will soon see, that turnover should be related to island area
(through effects of area on extinction rates) and/or isolation (through effects
on immigration rates. Neither was the case; instead turnover was approximately
inversely proportional to the number of species present. That is not forecast
by the model.
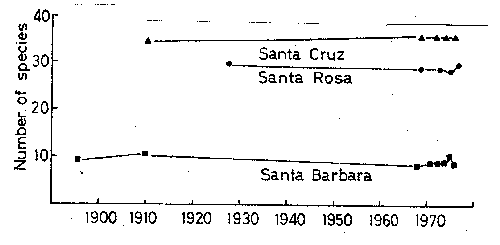
Figure
- The number of species in censuses of 3 of the California Channel Islands.
Figure
- % turnover in species numbers on California Channel Islands. (a) for nine of the islands. (b) for Anacapa as a function of time between pairs of surveys.
Why
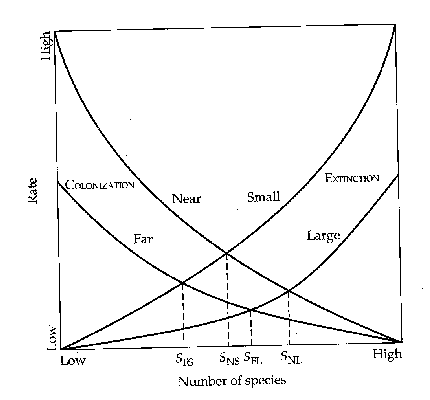
should turnover be related to island area or isolation? Consider first 2
islands at equal distance from the source, but differing in area. Long distance
(jump) dispersal is generally assumed to be a chance event, not directed or
goal oriented. In that case, dispersal probabilities and immigration rates onto
the 2 islands should be the same. Area, however, does affect the extinction
rate of colonists. The larger island should have 1) higher habitat
heterogeneity, 2) decreased intensity of interactions due to reduced niche
overlaps resulting from habitat heterogeneity and 3) larger population sizes
making chance extinctions less likely.
These
factors should be operative, at least in a relative way, independent of the
number of species present. Therefore, the extinction curves should have similar
shape, but have lower values for the larger island. Putting this comparison on
a graph, but using a linearized version of
immigration and extinction curves, we find a larger equilibrium number of
species on the larger island, but also a lower turnover rate on that island.
To
assess the effects of isolation consider 2 islands of equal size, but located
at differing distances from the source. With identical sizes we assume that
habitat heterogeneity, population sizes and interactions on the islands are
quantitatively identical, and thus they have the same extinction rate curves.
Immigration rates onto the more distant island should, however, be lower at any S since the probability of a successful
dispersal decreases (possibly exponentially) with distance. We can go further,
and suggest that the decrease should be most noticeable for species which tend
to be among the first colonists. Later immigrants with lower dispersal
capacities have only a slim chance anyways, and depend on rare, special
conditions like storms for successful immigration. For these species a change
in distance should mean less in shifting immigration rates. Once more we turn
these suggestions into a comparison on the graph. The more distant island has a
lower equilibrium number of species, but also a lower turnover rate at
equilibrium than an island closer to the source.
Figure
- Multiple immigration and extinction curves indicating effects of differences
in size and isolation on equilibria and turnover
rates. Brown and Gibson (1983).
These
comparisons can be combined in various interesting and complicated ways. Rather
than document the possibilities, it is probably more valuable to attempt to
list the assumptions and predictions of the basic MacArthur-Wilson model. Some
of the ideas in this list will not be fully examined until later in this
section.
Under What Conditions Does the Model Apply?
1) Islands are real isolates (rescue
effect, discussed below, not important)
2) Islands have comparable habitat heterogeneity
(complexity). There are no gross environmental changes over the time period of
colonization
3) Species counted on islands are
residents
4) There is a definable mainland species
pool
What Are the Characteristics of the Equilibrium?
1) It is dynamic
2) It is approached asymptotically
3) The process is inherently stochastic
4) The model and the equilibrium are
describing processes in ecological time
What Are the Characteristics of Turnover?
1) The process is not successional
2) Species replacements occur frequently
3) Immigration rates decrease with
increasing species numbers. Extinction rates increase with increasing species
numbers
What Influences the Equilibrium Number of Species?
1) Influenced by area through extinction
rates
2) Influenced by isolation through
immigration rates
3) Varies faster with area on distant
islands (see below)
4) Varies faster with isolation on small
islands (see below)
With this summary in mind, we return to
problems. With regard to Diamond's data, no combination of size and isolation
leads to the prediction that turnover rate is inversely (or in any other sense)
proportional to the number of species on an island.
Since
the data are repeatedly cited and classic, it's worth trying to understand why
this anomalous result was reported. There are a number of possible answers, and
arguments in the literature could be described by indicating that 'the fur has
definitely flown'. For one thing, the interval between the censuses was very
long. That may have had significant effect on the measured turnover. If the
time interval is long enough it becomes likely that some of the species which
had gone extinct at some time between the censuses also re-immigrated during
that interval (or the converse). In either case the measured turnover would
underestimate actual rates. To attempt to correct for that possibility, Diamond
and his collaborators went back to the Channel Islands annually during the
early 1970's, and also used thorough data gathered for Farnes
Island off Great Britain. The result of differences in the interval between
censuses is evident in Fig.8 (and reported in Diamond and May 1978). The result
for the Farne Islands is parallel. In either case the
apparent turnover decreases rapidly as the census interval increases. To show
you why, consider what happened to the meadow pipit on Farnes
between 1946 and 1974 (May and Diamond 1977). The pipit bred for 2 years, went
extinct in the 3rd, then went through 5 more cycles of
immigration and extinction over the remainder of the period. From annual census
records that indicates 11 turnover events in 29 years, where a census after 30
years would have recognized only a single extinction, as well as a constant diversity
of 6 species on the island. The same basic pattern applies to the Channel
Islands. Instead of turnover rates ranging from 17-62% (or .34-1.24% per year),
annual censuses indicate actual turnover rates of 1-10% per year, and are about
an order of magnitude larger than indicated by to 51 year interval for most
islands.
That's not the only corrective surgery which
has been suggested for the theory. It is also evident that monotonic rate functions
(particularly the immigration rate curve) may be overly simplistic. That should
be evident by drawing a parallel between accumulation of species on an island
and primary succession. When an island is newly formed (frequently volcanic) it
has no organic content in (and frequently no) mineral soil. The first plants
must be special sorts that have no requirement for nutrients from the soil (or
possibly no requirement for soil at all); instead they are soil formers,
leaving behind their nutrients extracted from the rock (as well as their
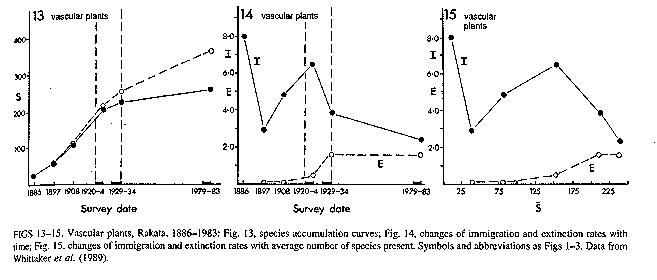
bodies) to improve conditions for later arrivals. Krakatoa,
East of Java, was not only a B movie, but a real historical event in the
1880's. What kind of immigration curve described the relationship between
immigration rate and the number of species on Krakatoa
after its formation. Depending on our definition of
immigration (does it end with landing on the island, or require initial growth
to be counted) and extinction (does a species have to reproduce at least once
on an island before we consider its loss an extinction?)
either immigration rate or extinction rate curves could be modified. For
simplicity we'll include the modifications in the immigration rate curve. Now it isn't monotonic decreasing; instead it
may have an initial rising phase representing the additional immigration
possible with the formation of soil. That is the naive logic. Reality isn't
quite so simple. Over the first 50-60 years since eruption (1883) the 'curves'
of the number of species accumulated over time for various plant groups seem
virtually straight lines; there is no decrease in rate of immigration over this
time. Similar arguments could be advanced, producing a similar curve shape,
with respect to other trophic levels. Equilibrium diversity and turnover rates
can be affected by these modifications.
Figure
- Colonization rates for vascular plants on Krakatau
measured in terms of total number of species (labeled 13), immigration and
extinction rates (14), and as functions of the number of species present (15). From Thornton et al. (1993).
Since
Diamond's data from the Channel Island studies suggested an appropriate
relationship between turnover and area-isolation effects, but could not
quantify them, an appropriate experimental test became important. Jim Brown and
his wife Astrid attempted such a test (Brown and Brown 1977). They studied the colonization of thistle
plants by assorted insects and spiders by repeated census following defaunation. Almost everything fit the basic
MacArthur-Wilson model, but... The turnover rate should be inversely related to
island distance (isolation) according to the theory. If we look at just
distance effects on the same island (i.e. area), the nearer island should have
a higher turnover rate. That's not what the Browns found. Instead, whether
plant 'islands' were large or small the turnover rates were higher on more
distant islands.
Site 1 Site
2
# of
plants Mean # species turnover rate # of plants Mean # species turnover rate
large- 16 3.82 0.67 9 5.25 0.29
near
large- 7 3.78 0.78 9 4.44 0.42
far
small- 56 1.89 0.78 21 2.21 0.69
near
small- 3 1.33 1.00 11 0.80 0.91
far
Based upon a 5 day census interval (remember
what Diamond's data ended up showing about the importance of census interval),
turnover rates were consistently higher on the more isolated plants. This
reversal of the expected pattern is explained as resulting from the effect of
repeated immigration of species onto near islands. The original model was
psychologically, if not explicitly, concerned with a degree of isolation which
made such repeated immigrations unlikely. Under real conditions repeated
immigration may be likely, particularly on near islands. Addition of a new
immigrant member into a small population reduces the probability of extinction.
This repeated immigration is, therefore, termed the 'rescue effect'. It could,
as I've just suggested, be presented as affecting immigration or extinction,
but since the key effect is on extinction rates, making extinction less likely
on near islands, that's where the curves are usually adjusted for rescue. When
the rescue effect occurs, turnover rates will tend to be directly proportional
to distance. The effect may be evident in data sets as divergent as these
studies of plant 'islands' and Diamond's New Guinea satellite island avifaunas.
Figure
- How the rescue effect modifies curves of immigration, extinction, and
turnover as a function of distance.
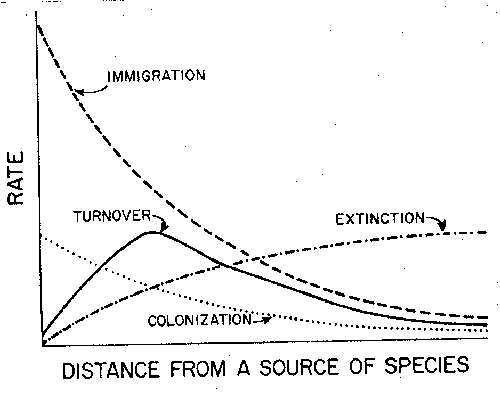
From
predictions of rescue effect occurrence we can make some general, graphical
predictions of how distance effects immigration, extinction and turnover. Note
that we are here graphing these rates against distance, not against species
numbers. The immigration rate declines exponentially with distance, as we've
previously seen in a variety of data. The extinction rate was previously
considered as depending on island area, and unrelated to distance; it would
have been a straight horizontal line on this graph before we considered rescue.
Now we recognize that the rescue effect bends the extinction curve down at low
distances. When we combine these curves to estimate turnover as a function of
distance, it has an intermediate peak; turnover is highest at those distances
when both immigration and extinction rates are high. Very close to the source
extinction rates are low; at large distances immigration rates are low.
The
next correction to the simple model is one which questions the validity of the
species-area relationship. Are projections of area-dependent extinction valid
for the total range of island areas studied? Of course, I wouldn't be suggesting
the question unless something were amiss. There is a
problem on very small islands. MacArthur and Wilson recognized that possibility
in presenting the basic theory, and suggested such islands were unstable,
should have very high turnover rates, and probably not have area-dependent
extinction curves. Basically, they thought any area effects would be masked by
the instability of the islands. The figure in the original monograph showed a
split curve for extinction rates, i.e. an unpredictable area effect. The
original data used to construct that graph was drawn from studies of the
ecology of the Kapingamarengi atoll system in the
Carolina Islands in Micronesia (Niering 1963). These
small outcrops appeared to show a threshold in the species-area relationship at
about 3.5 acres in area. It was suggested that the instability was not habitat
destruction by physical forces, but instability in the presence of fresh water.
Below the threshold area the water table on the island is saline; fresh water
availability depends totally on frequent rains. Above the threshold area there
is a permanent 'lens' of fresh water, and extinction
of plant and animal species dependent on fresh water becomes much less likely.
Figure
- Predictions of extinction rate characteristics on very small islands. From
MacArthur and Wilson (1967)
Modeling Effects of Disturbance on the Equilibrium
Theory
It is apparent that disturbance can have
important consequences for observed equilibria or the
lack thereof. What is difficult is the fact that disturbance has effects on the
survival and/or reproductive success of individuals. A disturbance, unless
extraordinarily massive, does not affect every member of a population. Modeling
on an individual basis has been difficult or impossible until recently. An
Italian group (Villa et al. 1992) attempted to evaluate the effect of regular
disturbance at differing intensities on the equilibrium. The following were
their conditions:
1) Island
habitats were equally distant from the 'source', but differed in size, from 50
'cells' (each cell was a potential site for an individual) to 1100.
2) There were
64 species. Each had its own mean lifespan, interval between reproduction, and
a range of clutch sizes from minimum to maximum, i.e. a life history. Each
species also has a relative dispersal capacity.
3) A
colonization species pool with relative abundances in the pool set.
4)
Colonization occurs by randomly allowing individuals to disperse according to a
negative exponential distribution (but distributions are affected by relative
dispersal distance). They are successful if they land on an empty cell. If so
their life histories determine whether the population grows or goes extinct.
5)
Disturbances occur periodically. Intensity varied from 0%-75%, where this
probability was applied to each individual, and determined the likelihood of
the individual being killed by the disturbance.
Some results of this simulation seem about as
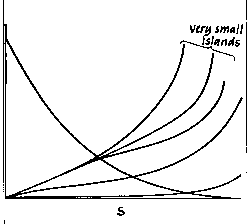
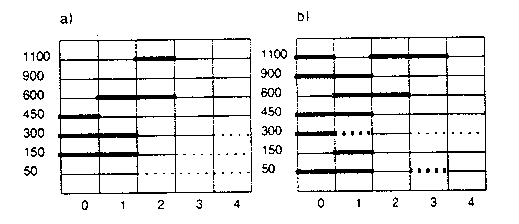
you might have predicted. Some are 'strange'. The two figures below show you
some of the key results. Figure 11 shows you their eyeball estimates of the
conditions which resulted in equilibrium. Over the 120 time intervals their simulation
ran, slow-growing organisms never reached equilibrium on large islands, but did
on small ones when there was no disturbance (indicated by 0 on the x-axis).
They could not reach equilibrium on any islands at higher levels of
disturbance. Fast growing organisms (on the right) could reach equilibrium on
any size island in the absence of disturbance, and with the larger population
size possible on very large islands, could even reach equilibrium in the face
of moderate disturbance levels (i.e. levels 2 and 3).
Figure
- indications of relative equilibrium (thick lines) in 10 simulation runs for
islands with different numbers of cells (the y-axis) and different intensities
of disturbance (the x-axis: 0-no disturbance, 1-10% effect, 2-25% effect, 3-40%
effect, 5-75% effect). Part a is for organisms with
'slow' growth (low clutch size, longer interval between reproduction, longer
lifespan) and part b for 'fast' growing individuals.
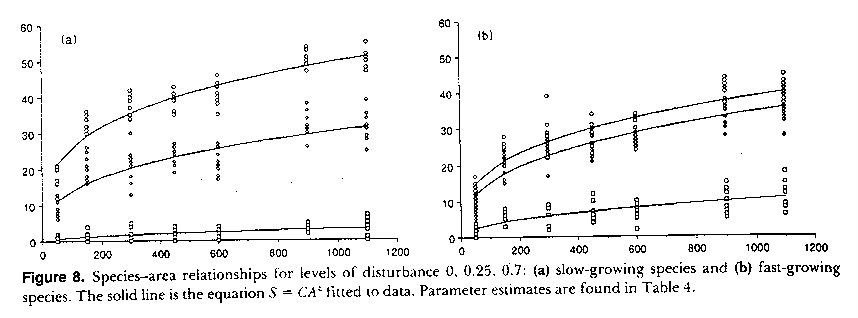
Figure
- Species-area relationships for a) slow-growing and b) fast-growing organisms
affected by disturbance. The 3 curves are for no disturbance (top curves), 25%
disturbance effect (middle curves) and 75% disturbance (bottom curves).
Evident
in the above Figure is the community level effect of disturbance. Disturbance
lowers the overall number of species resident, but if there is sufficient time
for equilibrium to have been reached, life history makes a difference. At low
levels of disturbance, the slow-growing species attain a higher diversity. At
high levels of disturbance, species are not able to remain around long, and a
greater diversity can be achieved by being a good colonizer (a weed, rapid
population growth, etc.). Counter-intuitively, when the actual fitting values
for these curves are assessed, the steepness of the species area curve
increases when disturbance is present, even though overall diversity decreases.
What all this tells us is we need to know more about the effects of disturbance
in real communities. The real world is affected by disturbance on a
more-or-less frequent basis, and conservation models based on an equilibrium
paradigm need to be re-considered to incorporate some indication of the effects
of disturbance.
References
Bially, A., and H. MacIsaac. 2000. Fouling mussels (Dreissena spp.)colonize benthic soft sediments in Lake erie
and facilitate benthic invertebrates. Freshwater
Biology 43:85-97.
Brown, J. and A. Brown. 1977. Turnover rates in insular biogeography: effect
of immigration on extinction. Ecology
58:445
Crawley, M.J. and J.E. Harral. 2001.
Scale dependence in plant biodiversity. Science
291:864-868.
Diamond, J. 1969. Avifaunal equilibria
and species turnover on the Channel Islands of California. Proceeding of the National
Academy of Science 64:57.
Diamond, J.M. 1973. Distributional ecology of New
Guinea birds. Science
179:759-69.
Diamond, J.M. and E. Mayr. 1976. The species-area relation for birds of the
Solomon archipelago. Proceedings of the
National Academy of Science 73:262-266.
Diamond, J. and R.M. May. 1977. Species turnover
rates on islands: dependence on census interval. Science 197:266-270.
Gilbert. 1980. The equilibrium theory of island
biogeography: fact or fiction?. Journal of Biogeography 7:209.
MacArthur, R.H. and E.O. Wilson. 1967. The Theory of Island Biogeography. Monographs in Population Biology, Princeton Univ.
May, R.M. 1975. Patterns of species abundance and
diversity. in M.L. Cody and J.M. Diamond (eds.)
Ecology and Evolution of Communities. Harvard Univ. Press,
Cambridge, MA. pp.81-120.
May, R.M. 1976. Patterns in multi-species
communities. Chap.8 in Theoretical Ecology - Principles
and Applications. R.M. May (ed.) Saunders, N.Y., N.Y.
p.151-157.
Niering, W.A. 1963. Terrestrial ecology of
Kapingamarangi Atoll, Caroline Islands. Ecological Monographs 33:131-160.
Power, D.M. 1972. Numbers of bird species on the
California Channel Islands. Evolution
26:451-463.
Simberloff, D. 1976. Experimental zoogeography of
islands: effects of island size. Ecology
57:629.
Thornton, I.W.B., R.A. Zan, and S. van Balen. 1993. Colonization of Rakata
(Krakatau Is.) by non-migrant land birds from 1883 to
1992 and implications for the value of island equilibrium theory. Journal of Biogeography 20:441-452.
Villa, F., O. Rossi, and F. Sartore.
1992. Understanding the role of chronic environmental disturbance in the
context of island biogeographic theory. Environmental
Management 16:653-666.
Whitehead, D.R. and
C.E. Jones. 1969. Small islands
and the equilibrium theory of island biogeography. Evolution 23:171.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}