Classical Species Radiations
Readings: Turelli, M. et al. 2001. Theory and
Speciation. TREE 16:330-343. hotlink to paper
Most
students of biology have become familiar with Charles Darwin and his work
with the famous finches of the Galapagos Islands that are named
after him. Indeed, much has been learned from this relatively small
group of birds, with important studies on this group of birds continuing
to this day by Peter & Rosemary Grant of Princeton University.
We
will explore outstanding examples of species radiations here, not
only because they give clear hints of how evolution and speciation occur,
but because these same taxa are in some cases threatened with extinction
as a result of man's activities. The examples we will consider are all
island faunas – Galapagos finches, Hawaiian honeycreepers, Hawaiian Drosophila and cichlid fishes of
the
great lakes of Africa. These groups of fauna are the
best documented examples of species radiations.
Galapagos Finches:
see the Web site: Voyage
of the Beagle to find out
more about Darwin's travels
aboard the Beagle).
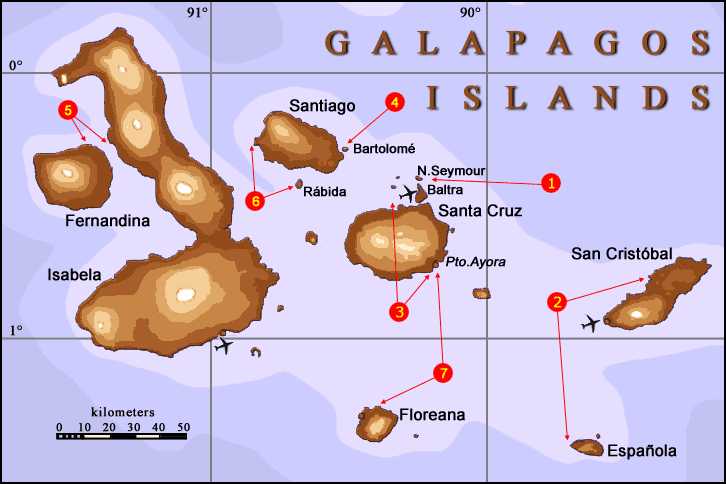
see image: Galapagos
Islands
The
Galapagos islands are a group of 16 major islands plus
numerous smaller islets (total = 32) located on the equator about 1000 km
west of the Ecuadorian (South American) mainland. The archipelago
was formed following volcanic eruptions of the seafloor ca. 3 - 5 million
years ago. The islands, because of their remoteness from mainland
sources of plants and animals, have a relatively impoverished biota.
Twenty-six species of land birds occurred naturally on the Galapagos
Islands prior to human introductions of others, and 13 of these are
finches. The islands also support 4 mockingbird, 2 flycatcher, 2
owls, 1 hawk, 1 dove, 1 cuckoo, 1 warbler and 1 martin species (Pianka 1983).
The
finches are thought to have evolved from 1 mainland species (or possibly a
Cocos Island species that reached the islands long ago. The island
finches belong to a distinct subfamily of finches, endemic to the Galapagos and
Cocos Island (Costa Rica). Cocos Island is several hundred km north
of the Galapagos and about the same distance from the mainland. It
supports only 1 finch species, the Cocos Island finch Pinaroloxias inornata. This finch species,
unlike those on the Galapagos, has had no opportunity for geographic isolation
(other than from the mainland). Thus, its gene pool has never been
split and additional species have never evolved there. The Cocos finch is a
generalist feeder, in contrast to many of those on the Galapagos which exhibit
specialist feeding niches.
On
the Galapagos Islands, adaptive radiation has resulted from
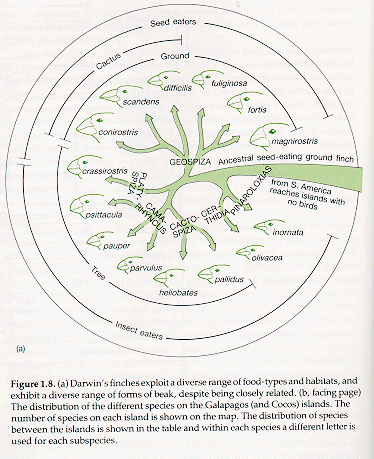
geographic isolation and reduced gene flow among islands. Three distinct
genera (Geospiza, Camarhynchus
and Certhidea) occur on the islands.
Members of these genera differ in where
they forage, how they forage, and what they eat. The ground-finches (Geospiza) include 6 ground-foraging species with
broad beaks that crush different species and/or sizes of seeds, or use
flowers of the Opuntia cactus. The
6 species of Camarhynchus finches forage in
trees, have narrower beaks, and eat either vegetation (1 species) or
different sizes of insects. C. pallidus
uses a stick or cactus spine to probe wood for insects. The other
genus has one species Certhidea olivacea, is called a 'warbler-finch'. It is
widespread on almost all of the islands and islets.
Other
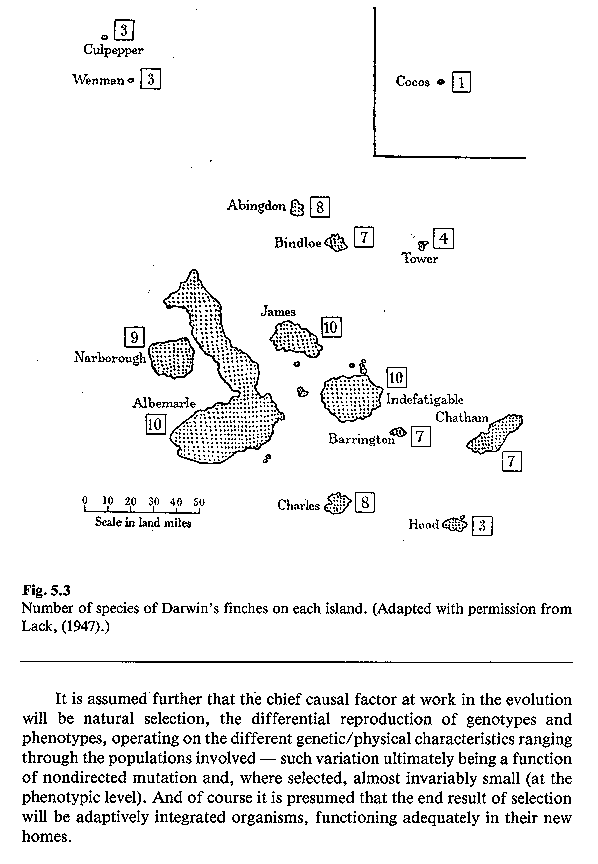
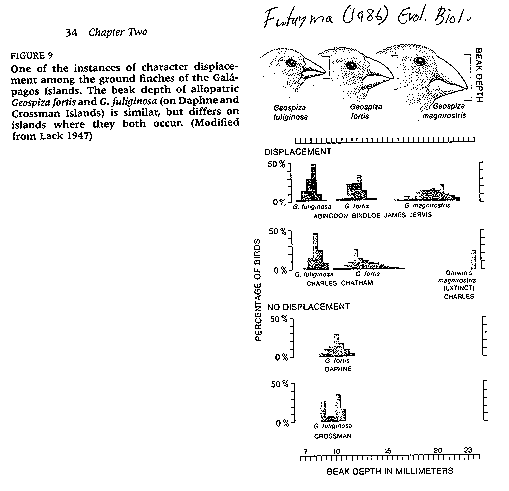
than Cocos Island, between 3 and 10 finch species occur on each island. see image:
finches (Ruse 1982). Beak lengths and depths vary widely
from island to island, and in some cases, provide limited evidence for
competitive displacement. For example, on islands (Abington, Bindloe, James, Jervis, Albemarle, Indefatigable,
Charles, Chatham) where Geospiza fuliginosa and G. fortis occur sympatrically, they tend
to have widely divergent beak depths.
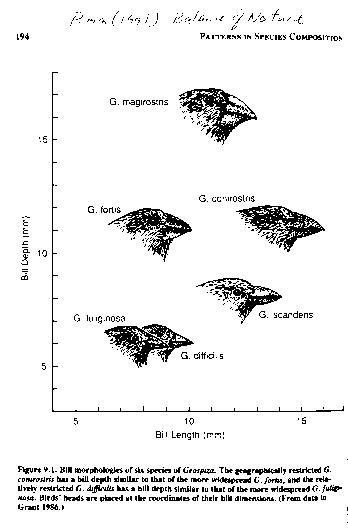
Because beak morphology is a critical feature in diet determination,
this separation is viewed as adaptive because it would tend to minimize
competition between the taxa during periods of resource scarcity. On
these islands G. fuliginosa
has a much smaller beak than co-occurring G.
fortis. However, only one of these
closely-related species is supported on the islands of Daphne
and Crossman. The beak morphology of these species are very similar to each other when they species occur in
isolation.
Many authors have cited this
as a classic example of character displacement, though before it can be
confirmed, 4 conditions must be met:
1) the change in mean character state (e.g. beak shape) in
areas of overlap should not be predictable from variation within
regions of overlap or areas of isolation;
To
date conditions 1 and 3 have been demonstrated. The variation observed in
G. fortis in isolation on Daphne greatly
exceeds that observed on other islands. Moreover, heritability of the
trait for beak morphology, which can range from 0 (no relation between parental
and offspring morphology) to 1 (absolute relationship) has been estimated as
0.82. Thus, there is good evidence that beak morphology is genetically
based. Criterion 2 cannot be met since there is only one Daphne and one
Crossman. Satisfying this requirement would necessitate observing the
same pattern on other islands like Daphne and Crossman. However, Peter
and Rosemary Grant recently demonstrated that co-occurring finches diverged in
beak morphology after a drought which greatly reduced seed abundance, relative
to their respective beak shapes prior to the drought. This provided good evidence in support of
condition 4. Divergence in beak shape
facilitated a divergence in diet and minimization of interspecific competition
during this critical period which John Wiens has
referred to as a 'resource crunch'.
Indeed, competition may be a very ephemeral or transient process that
may occur only sporadically, but its effects may be both long-lasting and very
significant.
Other
interesting patterns have been reported for the Galapagos Geospiza
finches. First, larger islands in the archipelago support more species than
smaller islands, likely owing to more habitats and more area of each
habitat. In addition, the number of finch species declines in relation to
the extent of isolation (= mean distance from
other islands) of the island relation to others in the group. This
concept will be explained in considering island biogeography. At the same time, the number of endemic
species increases with the degree of isolation.
Two
closely related taxa, G. fortis and G. conirostris, have completely different distribution
patterns and never co-occur. The same
pattern holds for G. difficilis and G. fuliginosa, though the patterns are not as clear
because they occasionally co-occur.
Grant and Schluter (1984) argued that G. conirostris
and G. difficilis are found on very few islands
because of interspecific competition. G.
conirostris is found on islands at different ends
of the archipelago, suggesting that it has had opportunity to colonize
islands in between. They attributed its rarity and absences on additional
islands to competition from other finch species.
The
pattern for G. difficilis is more
compelling. It is found on 4 widely separated islands and extinct on
4 others. This species co-occurs with G. fuliginosa
on large central islands (presumably with large habitat area to support
large populations), but do not co-occur on smaller central or small outlying
islands. Grant and Schluter used statistical
comparisons to suggest that in both cases the species co-occur much less
than one would expect by chance alone.
Dan
Simberloff (1984) argued that Grant and Schluter had
made incorrect statistical comparisons and that available evidence did not
support the competition hypothesis. Specifically, he stated that Grant and Schluter's method of assuming all 6 species were equally
likely to co-occur (their null hypothesis) was inappropriate since these two
species (G. conirostris and G. difficilis) were so rare. He argued that these
species overlap with few species precisely because they are so rare that they
cannot co-occur with other species.
It is widely believed that Darwin's finches
evolved from a single ancestor, possibly the Cocos finch. The original
colonists to the Galapagos island experienced adaptive radiation, a process
whereby the species diversified to exploit a wide variety of available (and in
this case, vacant) niches. As in the following cases of the Hawaiian
honeycreepers and cichlid fishes of the African Great Lakes, closely related
taxa co-exist by exploiting different habitats (trees vs. ground) or food
sources (eg. insects vs. seeds). If individuals with
similar niches practiced assortative mating, in which they preferentially mate
with individuals sharing similar traits or habits, then sympatric speciation is
possible (assuming that the behavior that separates sub-species is
heritable). In each of these cases of species radiations, the
diversification of the lineage occurred in an environment of low native species
diversity at the time of invasion - in other words, it appears that
diversification of the lineage was not constrained or prevented by pre-existing
competitors.
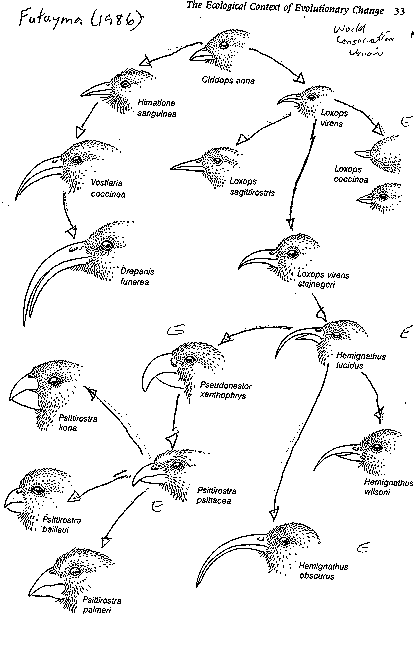
see image: honeycreeper - Futuyma 1986 (birds marked
with an 'E' are endangered or extinct).
The
honeycreepers of Hawaii are another remarkably speciose
group of birds; in fact, these birds experienced much greater adaptive
radiation than Darwin's finches, though sadly, many of the taxa have been
driven extinct owing to introduction of diseases, other passerine birds and
mammals, and destruction of habitat. For example, on Laysan
Island in the northwest region of the Hawaiian chain, introduced rabbits and a
large windstorm destroyed vegetation that resulted in the extinction of the Laysan honeycreeper. As well, bird pox virus and
avian malaria, the latter of which was introduced to the Hawaiian islands by mosquitoes on ships in 1826 have been attributed
with sharp declines in Drepanidid species diversity.
Currently, honeycreepers occur primarily at altitudes above 600 m on the main
islands and on several smaller remote islands in the NW part of the
archipelago. Mosquitoes, by contrast, occur primarily below 600 m, and
overlap very little with the honeycreepers. Many of the native birds are not
found at low latitudes because of habitat conversion (e.g. golf courses, sugar
cane). However, at the lower altitudinal end of the honeycreeper species
ranges, between 2 and 7% of individuals have avian malaria. This
frequency is 29% in one species that is highly mobile. Honeycreepers from
remote islands in the chain (which were less likely to ever come in contact
with introduced diseases) were highly vulnerable
to malarial infection, whereas some infected birds in highly populated regions
were apparently very healthy. Thus, it is not clear exactly what role, if any,
diseases have played in these extinctions.
Rats
have also played a large role in species extinctions. It is believed that rats
were introduced early on during human colonization of the islands (by
Polynesians 1000 years ago) and that many (~50%) native birds went extinct at
this time; 50% of the remaining species went extinct following European
colonization, and 50% of these remaining species are currently endangered.
Nine
extinctions have occurred since introduction of malaria to the islands.
Of these, 6 occurred on Lana'i, Moloka'i
and O'ahu; however, these small islands have been
radically modified by humans, thus habitat destruction appears to have played a
large role. Stuart Pimm has suggested that behaviour may have affected 2
large, nectar-feeding drepanididae species that have
been driven extinct and two smaller ones that have been driven to extirpation
('akoekoe and 'i'iwi) on
some islands. The large species were very aggressive
and were capable of defending high quality resources, but were poorly
adapted to exploit poor quality resources. He speculated that human
development reduced the high quality resources (e.g. native hibiscus) and that
the larger species were more vulnerable to extinction than smaller taxa that
were capable of exploiting the poorer quality resources that remained (e.g. 'akoekoe and 'i'iwi).
Another species, the rare 'akiapola'au, a specialized
insectivore, has become endangered because it lives primarily in large koa trees, forests of which have been felled as a wood
source and for tourist toys. Another endangered species, the palila, a granivore, exploits
seeds of one tree, the mamane. This tree has
been adversely affected by introduced goats and sheep, thereby endangering the
bird.
Thus,
it appears that many of the honeycreepers are endangered because of their
extreme specialization (habitat or food), which itself is a result of dramatic
adaptive radiation. This problem may be compounded by introduced
diseases, mammals, and exotic birds.
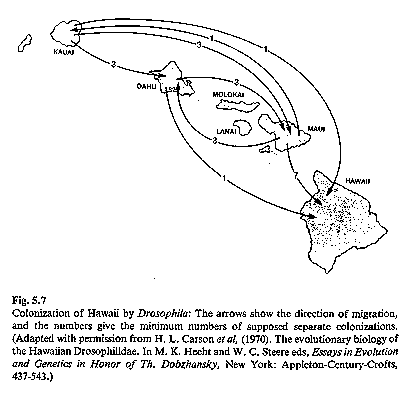
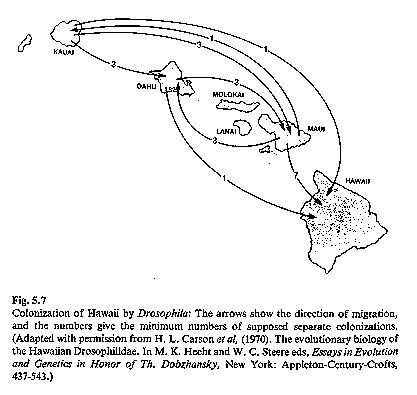
Hawaiian Drosophila
see image: Drosophila phylogeny of picture-wing Drosophila
of Hawai'i, representing
only a small fraction of all flies on the islands. (Futuyma 1986)
The Drosophila (fruit flies) of Hawai'i consist
of nearly 700 different
species, each typically restricted to one island. The islands vary
tremendously in age, Hawai'i being the youngest (~400,000)
years old); even this island has endemic species. Island age
increases progressively to the northwest including submerged
seamounts in the Pacific off the Asian coast. Speciation of
these flies is also speculated to have occurred as a result of
adaptive radiation. The speciation mechanism is not entirely
clear, though two main models are:
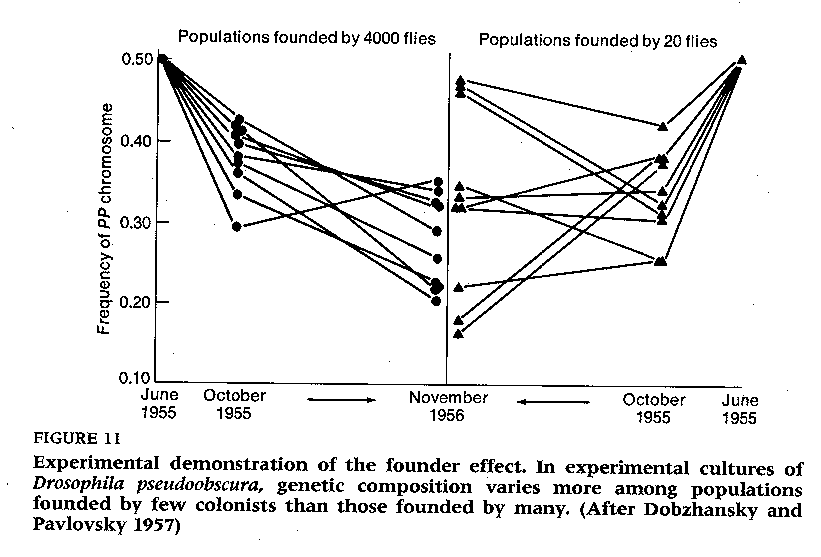
1) Mayr's founder effect
model: a few new colonists to an island represent only a small fraction of the
total gene pool. Genetic drift then results in dramatic 'peak shifts' in
the genome of colonizing species relative to the parental stock so long as gene
exchange is precluded. This genetic change in the population would occur
very rapidly and would be driven by local selective pressures in a relatively
homogeneous environment. (This idea formed the basis for the idea of
punctuated equilibrium model).
see founder effect with flies (Futuyma 1986)
Hampton
Carson modified this model slightly to suggest that certain loci form strong epistatic relationships (synergistic effects caused by
multiple loci) and a 'closed variability system'. For example, in fruit flies,
genes controlling mate recognition and behaviour may form such a closed
variability system' in which females will recognize only specific behaviours or morphologies when preparing to mate (=sexual
selection). Carson believes that these blocks of genes become destabilized
during founding events as the new population experienced exponential growth in
an altered selection environment. Recombinants that previously had low fitness
may now thrive, bringing the new population to a new
'adaptive peak'.
Templeton
modified this model suggested that change in allelic frequencies for a few key
genes (owing to genetic drift) could precipitate changes in modifier loci in
the new environment leading to a new co-adapted state of the character.
For example, Ken Kaneshiro has determined that some
species differ only by one mutation; this mutation modified the behavioural repertoire of males during mating
rituals. Derived (=new) species females would recognize the ritual and
mate with the individual whereas ancestral females (parental species) would not
and the species were behaviourally (sexually)
isolated from one another.
2) Brian Charlesworth has
argued that founding may not be the impetus for speciation, but that it may
occur as a result of adaptive divergence from the parental stock under a new
regime of selective pressures. Many studies have illustrated that
isolating mechanisms are determined by numerous genes. It is not clear
whether this can be applied to the Hawaiian Drosophila, as this view
maintains that speciation occurs as a result of gradual, sequential allele
substitutions.
An
interesting problem developed when immunological tests with Drosophila
suggested that the flies were far older than the islands. If this
were so, how could all of these endemic species occur on the
islands? Where did they come from? Dating of rock from each of the
islands and from the seamounts west of the islands have demonstrated that
island age increases east to west and that the oldest seamounts are older than
the flies. Thus, it appears likely that the ancestral flies once occupied
the seamount (when it was above sea level) and that they island-hopped to the
newly formed Hawaiian islands (which are located along a zone of crustal weakness; it is believe the islands were
formed and moved west on their plate), undergoing adaptive radiation as they
exploited new islands and habitats.
see image: leapfrogging
Hawai'i (Ruse 1992)
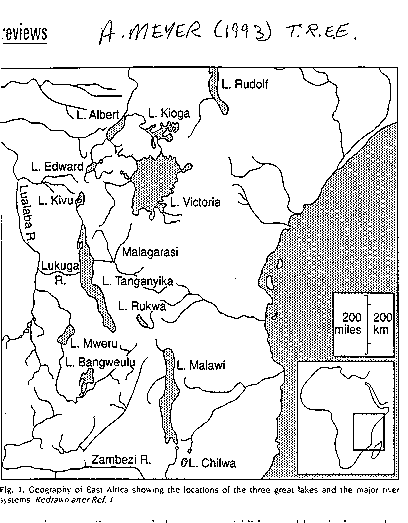
Cichlid
Fishes of the Great Lakes of Africa
Picture: cichlid
fish
The
great lakes of east Africa - Lakes Victoria, Malawi and Tanganyika are far
older than the North American Great Lakes
(see image: lake statistics - Lowe-McConnell 1994).
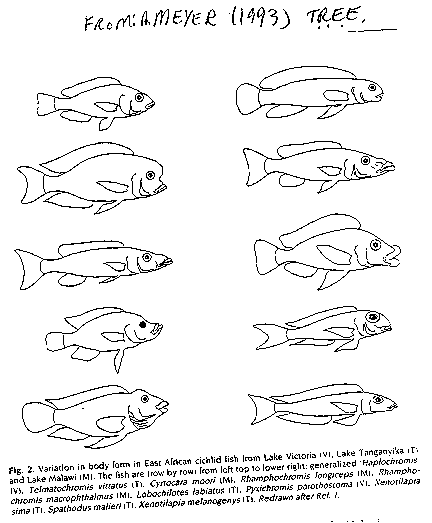
Many of these species are
highly specialized to exploit a specific habitat and food sources.
(see image: fish morphology - Meyer 1993).
For
example, in Lake Malawi, different cichlids feed on molluscs,
plankton, insects, benthic algae, and other fishes. The mouth parts
of these closely related species are often completely different, though
highly adapted for their particular form of feeding. Two genera feed
exclusively on scales of other fish, while one species feeds on other
fish's eyes. Indeed, if it was not known how closely related these
taxa are, they would almost certainly be assigned to different
families. The species have undergone dramatic adaptive radiation,
facilitated by the paucity of other fishes exploiting particular habitats.
As with the Hawaiian Drosophila, these
species have complex mating and behavioural rituals,
suggesting that sexual selection is an important factor isolating some
species.
The lakes:
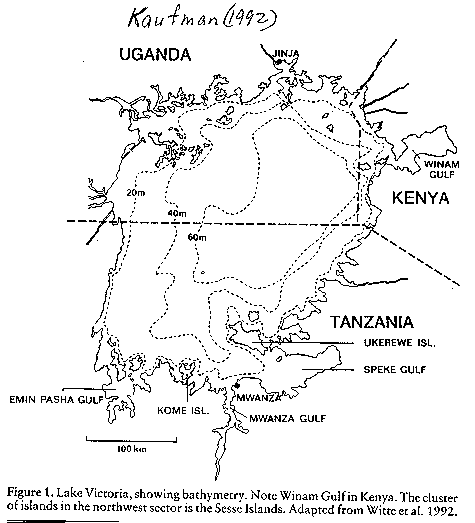
Lake Victoria: (see image:
lake morphometry - Kaufman 1992) the largest tropical lake in the
world (69,000 km2)with a maximum depth
of 84 m. The lake formed between 250,000 and 750,000 years
ago. There is some evidence that the lake level dropped drastically about
14,000 years ago, precipitating many extinctions but also permitting
allopatric speciation to occur. Indeed, Johnson et al. (1996) have
used seismic profiles to suggest that the lake was completely dried up
before 12,400 years ago; this would indicate very, very rapid evolution of the
cichlid assemblages in the lake. Coastal population growth with attendant
inflows of nutrients has caused exceptional cultural eutrophication problems.
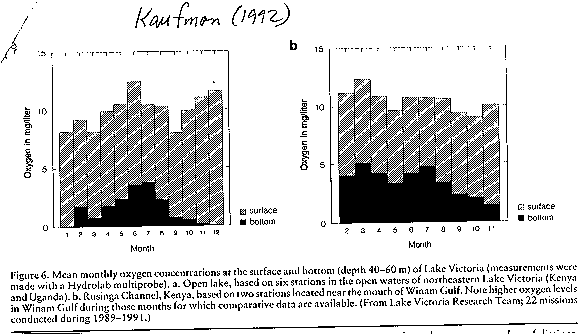
While the lake was once seasonally oxygenated throughout the water column,
it is now permanently anoxic below 20-30 m.
(see image: lake oxygen levels)(Kaufman
1992)
This dramatic increase in
anoxia has drastically reduced suitable fish habitat. As well, the
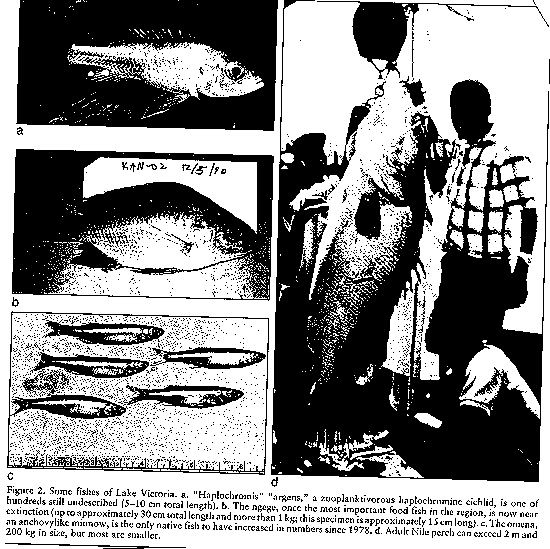
Nile perch (Lates niloticus)
was introduced to the lake, ironically to improve fisheries.
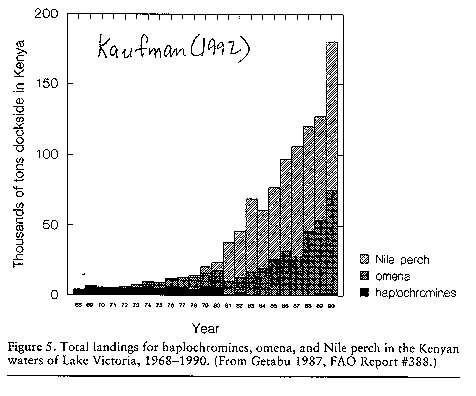
(see image: big fish - little fish)(Kaufman 1992).
The fish has caused
the extinction of native cichlid fishes and has reduced the fishery of
these and other native fishes (Kaufman 1992)
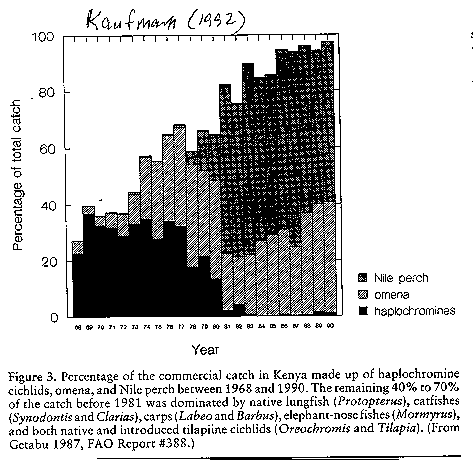
(see image: percent fish catch).
The lake had as many
as 300 cichlid species and 338 total species earlier this century (Worthington
and Lowe-McConnell 1994). These cichlids had radiated to occupy the
many feeding and habitat niches available in the lake. Sadly, as many as 200 of these cichlid species are now extinct
owing to the water quality changes and effects of Nile perch. Historically,
the lake supported a very large fishery (5x larger than any other African
lake) based on these cichlids (=haplochromines).
This fishery was based on many different species including Bagrus catfish,
lungfish, mormyrids and many others; today this
fishery has been virtually destroyed, and in its place is a fishery
dominated by Nile perch, Oreochromis niloticus and the endemic pelagic cyprinid Rastrineobola argentea.
(see image: total fish catch)(see
Kaufman 1992)
The fish community in Lake Victoria was also affected by intensive harvesting using increasingly small-mesh gill nets during the 1960's and 1970's. Declining catches prompted fish managers to introduce many tilapia species from adjacent areas; these species replaced the native tilapia species in lake (Worthington and Lowe-McConnell 1994). Nile perch were most likely introduced clandestinely as early as 1954, but were not observed until 1960. They were stocked into the lake in 1962 to improve the fishery by preying on the unused cichlid fishes (or so thought the managers); these fish could be harvested and, at the same time, provide hours of enjoyment for fishermen. As with many other invader species, the population grew very slowly but gradually until about 1980 when populations exploded. These fish have, ironically, proved a problem for 2 other reasons:
1) they
blow right through the weak nets used by fishermen;
2) they
are a lower quality fish than the natives they replaced.
At
present, the lake is in grave danger; eutrophication occurs unabated owing to
large nutrient inflows from the large human population living on the lake
banks. As well, increasing land-use in the basin around the lake has
resulted in loss of riparian vegetation. Deforestation has exacerbated
direct nutrient increases to the lake since this plant material previously
acted as a biological filter removing nutrients before the water entered
the lake. Finally, and perhaps most importantly, the lake has been invaded
by aquatic plants that forms a nearly continuous surface mat in shallow regions
of the lake. These plants strongly change light penetration, water quality and
habitat for native fishes and other organisms.
Lake Malawi
Like
its larger cousin Lake Tanganyika, Lake Malawi was formed millions
of years ago following faulting of the earth's crust. This faulting activity
resulted in deep fissures - the present day lakes. Lake Malawi is
the 3rd largest African lake but the second deepest. This lake has
defined seasonal changes, but is permanently stratified and anoxic below
250m (Lowe-McConnell 1993). The SE and SW sections of the lake have a
sandy bottom and support commercial fisheries of tilapia and
cichlids. The other regions of the lake are steep and rocky,
and support vastly different types of fish than sandy bottom areas.
The rocky areas support ~200 species of 'mbuna'
cichlids. These fishes are highly specialized and restricted to small patches
of rocky shore. They are separated from other taxa by stretches of
sandy shore or open water.
The
lake receded on 2 recent occasions - 25,000 and 10,740 years
before present; lake levels dropped dramatically ~1150 AD. and again between 1500 and 1850 AD. Malawi is thought
to have supported c.a. 545 fish species, of which 500 were cichlids.
Many of the cichlids apparently radiated because of their localized distributions and specialized diet - water level
fluctuations are thought to have isolated different groups, resulting in
micro-allopatric speciation. Many
non-cichlid fishes occupy the sand and mud areas (200 species).
Little is known about this group of fish. Because
of the basin's steep morphometry, its fishery
production has never compared to that of Lake Victoria, (a situation
similar to a comparison of Lake Erie vs. Lake
Superior). As with Lake Victoria, it is believed that the cichlid
fishes evolved very rapidly through adaptive radiation.
Intensive
fishing pressure in the lake into which Malawi flows (Malombe) resulted
in a collapse of the fishery there. In Lake Malawi, some fisheries have suffered
from deforestation and its effects on breeding streams, though fortunately,
exotic species play a very minor role in this system. Suggestions to
stock the lake with fishes from Lake Tanganyika were rebuffed and Nile
perch has never been stocked. The lake and its biodiversity is recognized as a World Heritage site. Despite this,
the rapidly increasing population of humans around the lake is imposing
tremendous fishing pressure, and is apparently responsible for the 'Utaka' (a schooling planktivore) and
predatory catfish decline (Lowe-McConnell 1996).
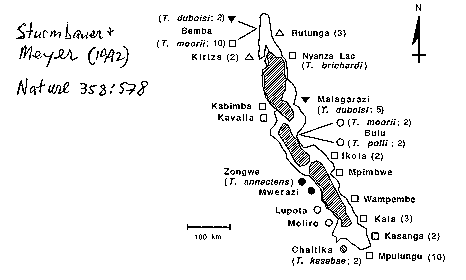
Lake Tanganyika (see
image: the big deep one)(Sturmburger
& Meyer 1992).
Lake Tanganyika is
the largest and deepest lake in Africa and the second deepest in the
world. It is also one of the oldest lakes in the world, aged between 9 and
12 million years old. The lake is permanently stratified and anoxic below
200 m. The lake supports 14 fish families and 12 tribes (flocks) of
cichlid fishes. Mouth-brooding cichlids occur only in this
lake. More than 220 species, mostly cichlids, inhabit rocky coastal
water, though open water fishes are also found. Most of the
deep-water fishes are poorly understood and little studied. In rocky
areas, cichlids occupy many feeding niches (algal-grazers, zoobenthic
feeders, omnivores, piscivores, plankton
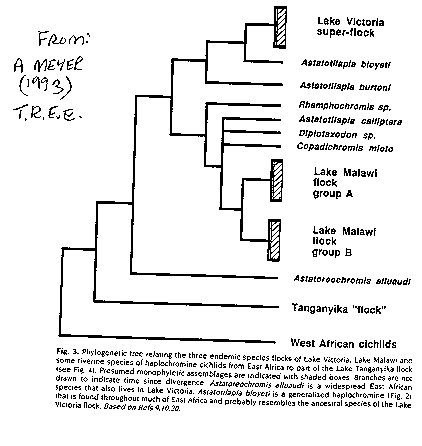
feeders). The cichlids belong to 49 endemic genera. Tribes are morphologically
and electrophoretically distinct and old, and can
be traced back to 7 ancestral lines. These fishes are also much
older than those in Malawi or Victoria (and thus are distinct from them).
(see image: phylogenetic tree of African cichlids)(Meyer
1993).
Studies by Sturmbauer and Meyer (1992) indicate that
the lake probably existed as three separate basins during dry
periods. As with Lake Malawi, it is thought that these drought
events tended to augment behavioural separation
of shoreline species, resulting in their divergence and speciation.
Many of the cichlids in Tanganyika (as in Malawi) are brightly coloured, raising the possibility that sexual or
social selection is an important component of their behaviour. The lake is
threatened by inflows of pollution, mainly silt-laden waters from rivers
owing to forest destruction in the watershed. Silt covers the
algae, the base of much of the foodweb and
alters fish spawning sites. Nutrients and industrial contaminants are a
growing problem. Because flushing time of the lake is exceptionally
long (thousands of years), any chemicals that enter the lake will cycle indefinetly (Lowe-McConnell 1996). Petro-chemicals
are one of the chemical groups most threatening to the lake. Fishing
pressure is also intense.
References
Begon, M, J. Harper and C. Townsend. 1986. Ecology. Sinauer,
Sunderland, MA.
Futumya, D.J. 1986. Evolutionary Biology.
2nd ed., Sinauer, Sunderland, MA.
Grant, P. and D. Schluter.
1984. In: D. Strong, D. Simberloff, L. Abele and A. Thistle (ed), Ecological communities: conceptual issues and the
evidence.p.201-233.
Johnson, T.C. et al. 1996. Late
Pleistocene desiccation of Lake Victoria and rapid evolution of cichlid fishes.
Science 273:1091-1093.
Kaufman, L. 1992. Catastrophic
change in species-rich freshwater ecosystems. Bioscience
42:846-858.
Lowe-McConnell, R. 1993. Fish faunas of the African Great Lakes: origins,
diversity and vulnerability. Conservation Biology7:634-643.
Lowe-McConnell, R. 1996. Fish communities in the African
Great Lakes. Environmental
Biology of Fishes 45:219-235.
Martens, K. 1997. Speciation in ancient lakes.
Trends in Ecology & Evolution 12:177-182.
Meyer, A. 1993. Phylogenetic relationships and
evolutionary processes in East African cichlid fishes. Trends in
Ecology and Evolution 8:279-284.
Pianka, E. 1983. Evolutionary Ecology.
Harper-Row, New York.
Pimm, S. 1991. The Balance of
Nature. University of Chicago Press.
Ruse, M. 1982. Darwinism defended. Addison-Wesley,
Don Mills.
Simberloff, D. 1984. In: D. Strong, D. Simberloff, L.
Abele and A. Thistle (eds.), Ecological communities: conceptual issues and
the evidence. pp. 234-253.
Sturmbauer, C. and A. Meyer. 1992. Genetic divergence, speciation and
morphological stasis in a lineage of African cichlid fishes. Nature
358:578-581.
Worthington, E. and R.
Lowe-McConnell. 1994. African
lakes reviewed: creation and destruction of biodiversity. Environmental
Conservation 21:199-213.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}