Biodiversity Patterns - RARITY
(see pg. 142-145 in text)
We
should recognize that the species approach, while it is inadequate on the larger
scale of conservation goals, can be important in identifying vulnerable
species. To this end, Deborah Rabinowitz (1981, 1986) described a general pattern of
species abundances, in which there were 7 different ways that species could be
rare as described by scientists studying them.
Rarity is in some of those cases, in the eye of the local beholder. Here
is a table, essentially duplicated from the text, indicating how species can
appear to be rare, be adapted to widely distributed low population densities,
etc.
Geographic range
Large Small Somewhere Common Locally Locally Locally large abundant over abundant in abundant in a a large range several specificPopulation in a specific habitats, but habitat, but
Size habitat type restricted restricted
geographically geographically Everywhere Constantly Constantly Constantly Constantly small sparse over sparse in a sparse and sparse and a large specific geographically geographically range and in habitat, but restricted in restricted in several over a large several a specific habitats range habitats habitat _____ __________ _____ __________ Broad Restricted Broad Restricted Habitat Specificity
Rabinowitz and her colleagues checked the patterns of
distribution and local abundance for the species sufficiently well described in
the flora of the British Isles. This flora has been studied and collected for
literally hundreds of years. It has also been carefully mapped. From maps of
individual species, it was possible to determine which species had large
geographic ranges and which small (N.B. within the British Isles). Global
species maps might alter the characterization of some species). From
descriptions of habitats where species had been collected, fellow scientists
were asked to decide whether the species were habitat specialists or
generalists, as well as whether species were at least somewhere locally
abundant. The kinds of descriptions are demonstrated below.
Descriptions:
Marshes, fens, heaths,
woodlands, and waste ground. A common weed of arable land.
– this is a description clearly for a habitat
generalist, at least some places locally abundant.
Restricted
to soil-filled crevices in scree slopes. – this is clearly the
description of a habitat specialist.
Scarce where present. Occurs as widely scattered individuals.
– this
is equally clearly the description of a species that is everywhere scarce.
The table of relative
occurrence frequencies for different kinds of rarity is shown below. There are
two numbers in each cell. The first includes all species for which sufficient
information was available, and for which there was general consensus among
observers. The numbers in parentheses reflect more conservative criteria for
inclusion, where 3 dissenting opinions or a consensus that the descriptions
were ambiguous for any trait resulted in exclusion of the species. Statistical
tests suggest that the two patterns ('liberal' and 'conservative' rules for
inclusion) do not differ significantly. Also interesting is a test that
indicates the three 'traits' are statistically independent. Think about that
one. There seem to be logical links among these traits, so that they should not
be independent. Common sense ecology would likely suggest that habitat
generalists ought to be widespread in distribution, but there are habitat
generalists with narrow (small) range. Logic that is not quite so strong might
also suggest that widespread species should be at least somewhere locally
abundant, but there are exceptions to this as well.
Geographic range Large SmallSomewhere large 58 (21) 71 (23) 6 (0) 14 (4) PopulationSize Everywhere 2 (0) 6 (0) 0 (0) 3 (1) small _____ __________ _____ __________ Broad Restricted Broad Restricted Habitat Specificity Thus, Rabinowitz
argued that species rarity could occur in three different ways:
1) restricted
geographic distribution;
2) narrow habitat
distribution;
3) low
local population abundance.
Of the British flora
analyzed, 39% had no component of rarity (i.e. they were not rare in any way),
while among the 61% that were rare in one or more ways:
59% had narrow habitat
specificity;
15% had small geographic
range;
7% had low population
size.
How one divides abundances
and ranges into abundant/rare, large and small affects one's
results. Pitman et al. (1999) analyzed trees in Peruvian Amazon
forest, and set 1 ind./ha as the abundance dividing
line, 1 vs. 2 or more forest type occurrences for habitat breadth, and a
political boundary (Madre de Dios) for
geographic distribution.
This analysis showed that
none of the trees were geographically confined (no small geographic ranges),
and only 13% were locally rare. The
rarest trees occurred at 1 stem per 36 Ha; over this forest this 'rare' species
would encompass 200,000 stems! One important consequence of this result
is, however, that conservation of truly rare species will require preservation
of a lot of land (Rickelfs 2000). Pitman
et al.'s results also differ markedly from the British forests surveyed by Rabinowitz. Much of this difference may relate to the
spatial scale used; consideration of spatial scale is obligatory when assessing
rarity. The lack of apparent habitat specialization by these tropical trees
runs in contrast to prevailing views. They also stand in contrast to Rapaport's Rules that high tropical
diversity is associated with habitat specialists and small geographic ranges.
Why is the separation
of forms of rarity important to conservation biology? Think about how the
limited resources available for conservation should be spent. Whole countries
identified as centers of diversity cannot be wholly conserved. Population,
development, and politics make any thoughts of that level of protection
ridiculous. Within such countries, what should be protected? Clearly, the best
approach is to preserve habitats and ecosystems. But which
ones? If information of this sort is available (or can be developed) for
species of various sorts (not just plants), then the form of rarity can be very
useful. A species that is rare due to limited geographic distribution, but has
fairly generalized habitat requirements and is locally abundant seems like a
good candidate for introduction into areas outside its current distribution
limits. A species with a wide distribution and high local abundance, but narrow
habitat requirements, probably cannot be successfully moved; it has likely
already sampled areas throughout its range. The likely solution is protection
of at least some of the local populations already established. Species that are
locally sparse, but widely distributed and generalized in habitat requirements
probably need no protection. In sum, species restricted to narrow niche
characteristics in two or more of the three traits are those most likely to
need some help. They can identify species, habitats, and communities on which
to spend our efforts.
There may exist biological reasons for species rarity. Kunin and Gaston (1993), in a review of rare and
common species, found that rare species (i.e. locally rare and geographically
restricted) differ from more common species. Rare species have lower
levels of self-incompatability, a greater tendency to
asexual reproduction, lower overall reproductive effort and poorer
dispersal abilities. In a sense, they tended to make the best of a bad
situation.
Groups
such as the World Conservation Union (IUCN) use indicators to determine which
species are rare and at risk of extinction. Programs can
then be developed for these species to alleviate their population persistence
problem. Typical indicators used for
this purpose include:
1) rarity;
2) rate
of decline (high rates being bad);
3) degree
of population fragmentation.
However, Hartley and Kunin (2003) have demonstrated that each of these 3
indicators is sensitive to the scale at which they are measured. Rarity can be measured using 3 metrics:
1) Extent of occurrence – essentially the total
range area (this tells us nothing about the distribution of individuals within
that range nor their abundance);
2) Area of Occupancy – typically measured by
summing occurrence of individuals in individual ‘cells’ of a grid
encompassing the entire range (again, telling us nothing about abundance at
each);
3) Population Counts – measured by population
census (highly specific population number for a specific location in the Area
of Occupancy:
Area of Occupancy (AOO)
___________________________________________________________
Population
Count (PC) Extent
of Occurrence (EOO)
(individuals) (range size)
ß------------------------------------------------------------------------à
fine scale coarse
scale
In the IUCN model, species
are considered critically endangered if the extent of occurrence is <10km2,
endangered if they occupy <500km2, and vulnerable if they occupy
<20,000km2 . A problem arises with the AOO criterion
however, since species mapped at high spatial resolution will appear rarer than
species mapped at coarser spatial resolution.
This means we are more likely to give lower extinction risk values to
species for which we have the least information (Hartley and Kunin 2003). In Britain, AOO is measured on a 100 km2
grid, meaning that species occurrences can only be mapped at no less than this
level of resolution (i.e. they cannot be called critically endangered since no
measures at the high spatial resolution scale are available).
Using measures of rate of
decline is also fraught with problems. As the term implies, rate of decline
will be measured by assessing population size over at least 2 points in time.
Four possible measures are possible using only range size and population number
(declines in range but not population number, declines in population number but
not range, both, neither). Field
evidence suggests that calculations based on decline at the coarse scale (AOO)
would yield lower decline rates than those calculated at a fine scale
(PC).
Third, the IUCN method also
considers population structure with respect to dispersal potential (i.e.
fragmentation extent). However, severe
fragmentation (many small, isolated populations) and lack of fragmentation (a
single or few subpopulations) can be considered as indicative of increase risk
of extinction. So, the relationship
between fragmentation and risk is not clear.
Thus all three criteria currently used by IUCN to designate species are
endangered or not as sensitive to spatial scale problems.
Hartley and Kunin (2003) note that 3 options exist for dealing with
spatial scale issues:
1) Use a standard spatial
scale to measure AOO – e.g. the 100 km2 as in Britain, even
though it has problems.
2) Measure each of the 3 metrics across as many
spatial scales as possible (basically Rabinowitz’s
approach). It overcomes the issue of
arbitrary scale size selection, which may or may not reveal true rarity. But it requires and produces a great deal of
information.
3) Combine information from multiple scales in a
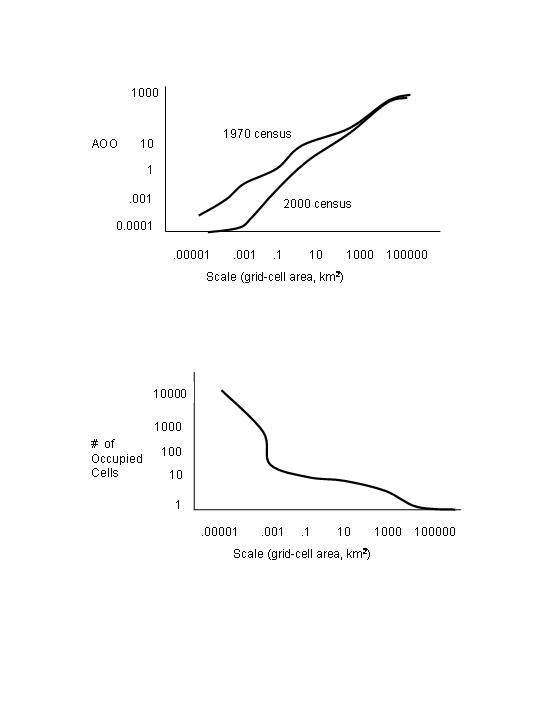
simple, unified, quantitative manner by plotting AOO vs. scale size.
Figure
4 – Hartley and Kunin (2003)
If we plot the AOO values
for more than 1 time period on the graph, we can determine whether the curves
have changed and whether the species is becoming more rare, and, if so, at what
spatial scale. If we use the multiscale approach, as
suggested by Hartley and Kunin (2003), we will be
able to calculate the value of each rarity metric and at any scale, and
therefore determine precisely how a species’ abundance and occurrence are
changing over time.
References
Harte, J. and E. Hoffman. 1989. Possible effects of acidic deposition on a Rocky
Mountain population of the tiger salamander Ambystoma tigrinum. Conservation Biology 3:149-158.
Hartley, S. and W. Kunin. 2003.
Scale dependency of rarity, extinction risk, and conservation priority. Conservation Biology 17: 1559-1570.
Kunin, W.E. and K.J. Gaston. 1993. The biology of rarity: patterns, causes and
consequences. TREE 8:298-301.
Pitman, N.C.A. et al. 1999. Tree
species distributions in an upper Amazonian forest. Ecology 80:2651-2661.
Rabinowitz, D. 1981. Seven forms of rarity. pp.
205-217 in The Biological aspects of rare plant conservation. Ed. by H. Synge. Wiley.
Rabinowitz, D., S. Cairns and T. Dillon. 1986. Seven forms of rarity and their frequency in
the flora of the British Isles. In M.E. Soule; (ed.)
Conservation Biology: The Science of Scarcity and Diversity. pp.
182-204. Sinauer.
Ricklefs, R.E. 2000. Rarity and diversity
in Amazonian forest trees. TREE
15:83-84.
{kind=link}