Conservation Biology Case Studies
text pp. 420-426,169, 432-441, 380-381,433, 553, 649, 83,
89, 288.
Conservation efforts
have classically been aimed at species rather than habitats, communities or
ecosystems. A number of studies may be identified which document
successful recovery of species from critically endangered status (e.g.
peregrine falcon, whooping crane), while others have proven either less
successful (black-footed ferret) or unsuccessful (passenger pigeon,
Pacific Island tree snail). Among the most controversial species currently
embroiled in conservation programs are the African cheetah (Acinonyx jubatus) and northern spotted Owl (Strix occidentalis caurina). We will review three cases
(black-footed ferret, cheetah, northern spotted owl) because they provide
interesting insights regarding conservation problems or because they involve
major controversies.
1) Black-footed ferret (Mustela nigripes): see www site listed on course homepage. Meffe
& Carroll (1997) pp. 83, 420-426.
This species is a
small member of the weasel family that formerly occupied plains and prairie
habitat from Saskatchewan to Texas. The species was listed as threatened
in the USA in 1967 and endangered in 1973. An initial recovery plan was
devised by the US Fish and Wildlife Service in 1978. By then, however, there were
no known wild black-footed ferrets. The species was re-discovered at a single
site in Wyoming in 1981. A Species Survival Plan (SSP) was developed based on
captive propagation of ferrets to eventually re-release ferrets into the wild.
Decline of the
species coincided with, and may have been caused by, the tremendous decline
(90-95%) in prairie dog abundance earlier this century. Prairie dogs are
the primary (90%) food of black footed ferrets. Prairie dogs were
targeted as pests because their burrows damaged farm implements and tractors.
In addition, the prairie dogs appear to have suffered from introduction of Sylvatic plague. A severe decline
in prairie dog abundance was evident by 1985, when ferrets were collected to begin
a captive propagation program. The ferret's behaviour and ecology had been
studied in the Wyoming population from 1981 to 1985.
Ferrets also suffered
from habitat fragmentation as the plains were developed for agricultural and
other purposes. By 1985, the species was limited to ~10 individuals in
two populations (South Dakota and Wyoming). One population (from
South Dakota) was placed in a captive breeding program without success
(Was this good conservation biology?). The other population was surveyed
but not captured; this population increased to 128 animals by 1984. This population
soon suffered, however, after plague was discovered among its prairie dog prey.
Ferret numbers appeared to decline sharply in 1985 despite treatment of 80,000
prairie dog burrows to control plague. Six ferrets were captured for a
captive breeding program but all died of canine distemper. Additional
collections of 6 and 12 animals were made for captive breeding programs.
The last free-ranging individuals died from distemper, resting the fate of the
species in the last 12 individuals in the captive breeding program. Goals were quickly set to maintain as much
(90%) genetic diversity as possible for a minimum of 50 years. Two
litters of kits were born in 1987, and by 1988 the first two isolated captive populations
were initiated (to minimize catastrophic extinction of the species).
Extensive research on
ferret reproductive biology, immunology and behaviour was conducted to maximize
offspring production and survival, and to prepare the animals for eventual
release in the wild. By the early 1990s the success of the captive
breeding program was evident in the birth annually of >100 kits. Ferrets
were reintroduced to southern Wyoming in 1991 with the permission and
cooperation of landowners (228 ferrets over the period 1991-4). Some
legal battles pitted ranchers' organizations and some environmental groups
against the Fish and Wildlife Service and its reintroduction plan. (A similar
problem arose with Florida panther captive breeding programs). Forty-nine
young individuals were given a soft-release (food provided at cage site after
the animals were exposed to the area [but still confined] for 10 days).
Some of these individuals (12%) survived the winter and reproduced
successfully. Ninety additional ferrets were released in 1992.
Coyotes appeared to be the primary predator and source of mortality, though
survival was moderate (20-25% for 30 days).
The ferret is still
in jeopardy owing to distemper virus, plague and low population numbers (genetic
bottleneck). However, the population is growing and introductions of
ferrets to a site in South Dakota (90 ferrets) and one in southern Montana (78
ferrets) in 1994-5 occurred. Care is exercised in selecting sites to minimize
the possibility of contact with plague (prairie dog prey) or canine distemper.
Local responses to ferret re-introduction are
also significant. The re-introduction sites are a first example of the problem
imposed by political opposition. The re-introduced ferrets are (in the language
of the U.S. Endangered Species legislation) a nonessential, experimental population.
Under this designation, the animals are protected at the re-introduction site,
but are left unprotected should they move into a farmer's field or a rancher's
pastureland.
2) African cheetah (see www site listed on course homepage)
In 1983, O'Brien and
colleagues reported that cheetahs had remarkably little genetic
variation. The species is limited principally to regions in sub-Saharan
Africa, though a small population remains in Iran as well. The population
is estimated to have declined by 50% in abundance (to ~10,000 to 20,000) by the
mid-1970s from the previous decade, largely as a result of habitat destruction
and hunting by humans. The population has continued to decline though accurate
estimates are not available of current population size. O'Brien et al.
[1983, 1985] speculate the total population is between 1500 and 25000
individuals).
In a study that
commenced the controversy over reasons for endangerment of the cheetah, O'Brien
and colleagues (1983) reported that 55 captive and wild-caught cheetahs derived
from two separate populations were monomorphic at all
47 allozymes surveyed. They compared their results with those for 43
fruit fly (Drosophila) species, 2
mouse (Mus)
populations, 1 cat (Felis)
population, and a large number of human (Homo)
populations. The cheetah had the lowest frequency of polymorphic loci
(0.0) and lowest average heterozygosity (0.0).
Overall, the cheetah had between 10 and 100 times less genetic variability than
other mammals.
See
cheetah genetic diversity O'Brien et al 1987)
O'Brien attributed
the patterns in cheetah to a severe population bottleneck followed by
inbreeding. The bottleneck would reduce genetic diversity as a result of
selection pressures and genetic drift. They attribute the bottleneck to
decimation of the population by legal and illegal hunting by African cattle farmers
about 100 cheetah generations ago. They suggested this genetic pattern
would also be consistent with a bottleneck 100 generations ago coupled with a
low population growth rate (which would allow drift to reduce diversity).
They suggest that current cheetah distribution is but a remnant of its once
global distribution (Africa, Asia, Europe, North
America), and that a low sperm count and abnormal sperm is evidence of a
bottleneck and inbreeding.
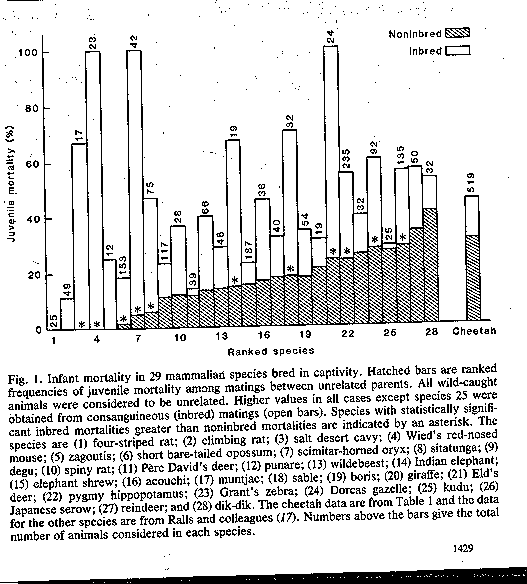
O'Brien et al. (1985)
also reported significant mortality of cheetahs in captive breeding programs in
zoos. Non-inbred cheetah mating had among the highest infant mortality
rates of all mammals surveyed. The same pattern was observed for inbred matings of cheetahs. As well, infant mortality rates
for inbred and non-inbred cheetah mating did not differ significantly, suggesting
that inbreeding has no pronounced effect today (largely because strong effects
were evident earlier).
See
mortality rates for mammals (O'Brien et al. 1985)
They also added 5
additional enzyme surveys to the previously studies 55 individuals, with the
same results. As well, 7 different skin grafts were performed on
non-inbred pairs of cheetahs (14 individuals) to determine whether different
cheetahs could serve as skin donors. Successful grafts depend on acceptance
of 'donor' tissue by the 'recipient' individual, which in turn is governed by a
group of genes called the 'major histocompatability
complex' (MHC). In all vertebrate, the MHC is the most polymorphic region
of the genome, thus it should be most useful in differentiating genetically
different individuals. All of the grafts succeeded through the typical
stage of rejection, though control grafts of house cat tissue were rejected;
thus, the cheetahs were incapable of immunologically
identifying other cheetah's tissues, though they did recognize cat
tissue. This suggested strong genetic relatedness of the cheetahs.
As well, an infectious feline virus wiped out a cheetah colony in captivity in
Oregon; O'Brien attributed the widespread success of the virus to genetic
uniformity of the cheetahs.
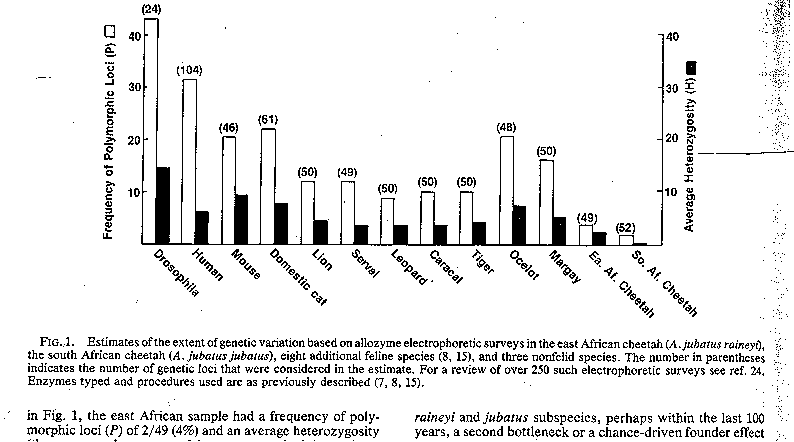
In a third study
O'Brien and colleagues (1987) again visited the cheetah issue to look at
frequency of enzyme polymorphisms and heterozygosity
levels in subspecies of the cheetah, the south African
form A. jubatus
jubatus and its east African relative A. jubatus raineyi.
Again they found very low levels of polymorphism (2 - 4%) and average heterozygosity (0.0004 - 0.014) in both groups; as well,
the genetic distance between subspecies was minimal (0.004) indicating that the
cheetah became genetically impoverished before the subspecies diverged.
They again stated that these genetic patterns are most consistent with 2
bottlenecks (one 10,000 years ago and another during the past century) followed
by inbreeding.
Menotti-Raymond and
O'Brien (1993) used two DNA analysis techniques - hypervariable
minisatellite loci and mitochondrial loci - to time
the bottleneck in the cheetah population. Genetic variation was observed
with these techniques. Based on expected mutation rates
and current levels of diversity, they back calculated the bottleneck to
between 3500 - 12,700 years and 28,000 - 36,000 years, respectively, for
mitochondria and minisatellite techniques. The
latter variable is likely an overestimate of the true time, though both
techniques place a bottleneck during the late Pleistocene (when many
extinctions occurred). These techniques also identified only 1 to 10% of
DNA diversity found among other out-crossed cat species.
More recently,
however, fireworks erupted regarding the cheetah's genetic and demographic status.
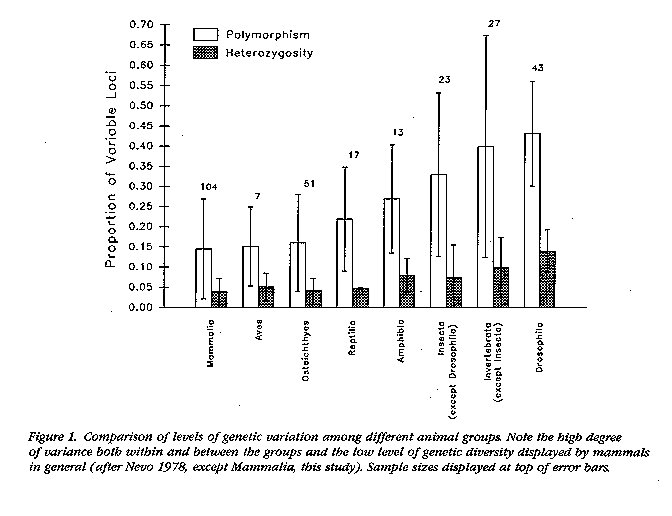
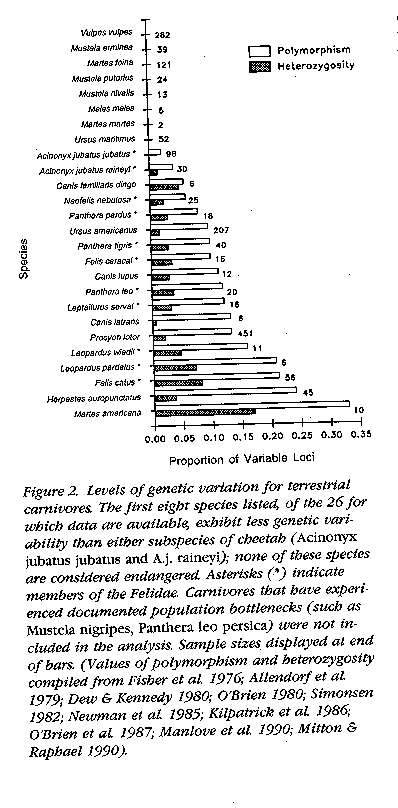
The lead argument was fired by Merola (1994) who
compared the cheetah's genetic variability with that of other carnivorous
vertebrates. She suggested that of 24 terrestrial carnivores surveyed, 8
had no heterozygosity (H = 0), while the remaining
ones averaged H = 0.042 (vs. H = 0.014 for the cheetah).
See
genetic structure of animals (Merola 1994)
See
Heterozygous Loci in Carnivores (Merola 1994)
She also stated that
the lack of breeding success and high infant mortality rates were due to poor
captive breeding program procedures, and that the feline virus that decimated
the Oregon cheetahs was effective because the cheetahs were held at very high
density. Cheetahs are solitary creatures in the wild and would therefore
rarely encounter densities like the Oregon situation.
She argued that
as long as recessive alleles (deleterious) were slowly purged from the
population, the resulting population could be relatively homozygous but without
inbreeding effects. The inbreeding
effects observed in cheetahs would thus be an artifact of the artificial
captive breeding environment.
Merola
acknowledged that the cheetah is suffering, but it is from a loss of habitat
and other adverse human effects. For example, habitat destruction has resulted
in population densities of one cheetah per 6 km2 rather than the old
rate of 1 per 100 km2. High
densities facilitate transmission and spread of disease and 'focusing' of
cheetah predators in the small reserves. Cheetah cub death rate is 93% in
the Serengeti reserves, of which 73% was attributed to predation by lions and
spotted hyenas (Laurenson et al. 1995). It
seemed as though reserves promoted predation because of the 'focusing' effect
of predators on remaining 'islands' of intact cheetah and predator habitat. Cheetahs are probably also suffering due to
food reductions associated with destruction of natural habitat used by the
cheetah's ungulate prey, and to hunting.
O'Brien (1994) fired
back in the same issue of Conservation Biology that Merola's
comparisons used only allozyme data for a relatively
small number of loci (< 22), while the cheetah allozymes data consisted of
52 loci. Thus, the cheetah studies used more loci but still found less
diversity. In addition, other more useful modes of genetic testing
(described above) yielded the same results - that the cheetah is genetically depauperate relative to other mammals, terrestrial
vertebrates and other cats. He also suggested that the high natural
mortality rate of cheetah cubs observed in Africa may have resulted from
discovery of dens (thus multiple killings of cubs rather than independent killings)
and to predators learning of den locations from researcher movements.
O'Brien does not preclude environmental problems as one of the issues
confronting survival of the cheetah, though he maintained that the larger
problem was genetically-based.
Laurenson
at al. (1995) flatly rejected O'Brien's notion that they led lions and other
predators to cheetah dens because they took precautions to prevent this
possibility. They also rejected the idea that predation rates were
inflated by predators killing all offspring in a den, because predation was a
major mortality factor for all dens, not just specific ones. They argued
that mortality of cheetah cubs in captivity was attributable to genetic factors
(<4%) and animal husbandry (78%). Thus, in the wild they suffer from
predation, while in captivity they suffer from poor breeding practices.
Robert May (1995)
reviewed the two sides and felt that there was evidence for genetic problems,
but that environmental effects were also very important. The issue is
unlikely to be resolved soon because of the entrenched views each side has adopted.
See Abstracts of:
1) Merola (1994)
1 and 2
The most recent
addition to the debate was the contribution of demographic modeling
contributed by Crooks et al. (1998). Using published
demographic data from the Serengeti. They found that the importance of
elevated cub mortality was relatively minor relative to the large effects
from variation in adult survivorship. This finding makes sense from
a 'demographic' perspective since the adults have high reproductive value and cubs low value. Further, they argue that focusing too
much attention on reducing cub mortality could be counteracted by small
increases in adult mortality. They summarize by stating that the
genetics vs. ecology debate is not helpful since both factors affect cubs
and/or adults.
3) Northern Spotted Owl (S. occidentalis caurina): Meffe
& Carroll pp. 380-381,433, 553, 649, 83, 89, 288.
Northern Spotted Owls
occur in the southwest region of British Columbia and in Oregon and
Washington. In all instances, the owl is rare (low abundance) even in the
best of habitats. In southwestern B.C., the owl was found at 14 sites,
with a total population of as few as 100 individuals (Dunbar et al. 1991). They attributed its rarity to habitat
destruction (logging, fires, development) and to Barred Owls which live in the same
old-growth habitat and which respond aggressively to spotted owl calls (thus
potentially limiting its habitat availability).
In the USA, the
northern spotted owl has pitted environmentalists against loggers to the point
where decisions regarding the fate of public lands have gone all the way to the
Supreme Court. (As an interesting aside, the case was resolved during
summer 1995 by the conservative-leaning Supreme Court in favour of preservation of essential lands for owl
habitat). The reason why this is such a contentious issue is that the
bird is heavily dependent on old-growth forests for nesting habitat, the same forest
currently under assault in the western USA and Canada. Because any decision
made by the U.S. Fish and Wildlife Service regarding preservation of old-growth
forests would be targeted legally by logging interests, the ecologists involved
knew that their science would have to stand up.
Tracts of forest
lands suitable to spotted owls in the western USA have declined at an alarming
rate.
See
Habitat Decline in USA (Meffe and Carroll 1994)
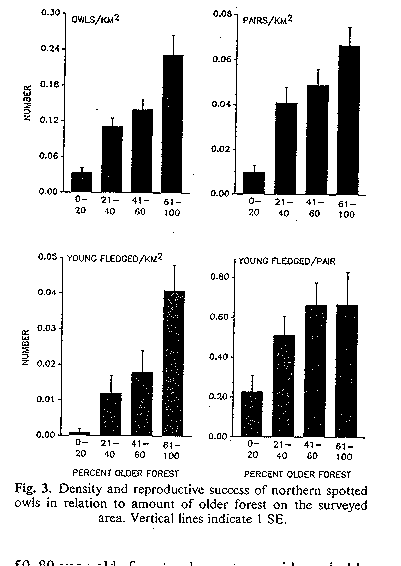
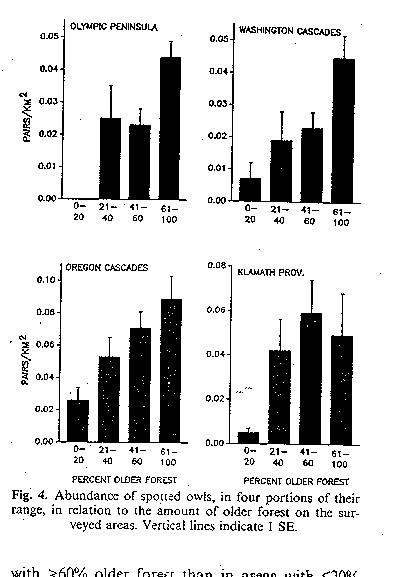
Bart and Forsman (1992) and Bart (1995) looked at spotted owl density
and breeding success in habitats of differing quality in Washington and
Oregon. In sum, the higher the percentage of old growth forest (good
habitat), the higher the owls/km2, breeding pairs/km2,
young fledged/km2, young fledged/km2, and adult survival.
See
owl density and success (Bart and Forsman 1992)
See
owl density (Bart and Forsman 1992)
See
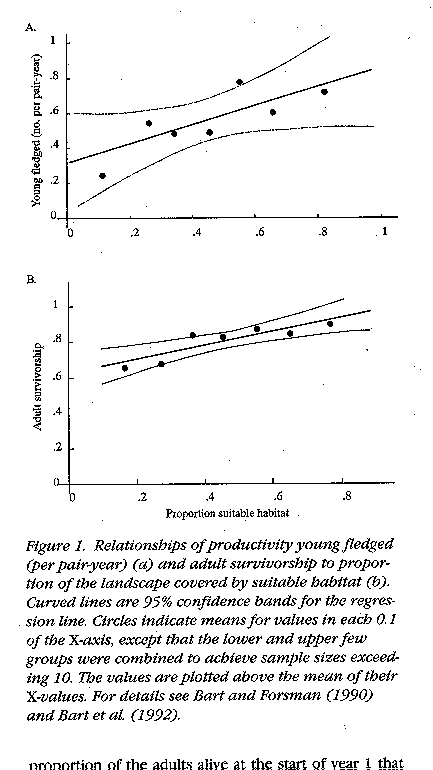
adult survival rate (Bart 1995)
In what is perhaps
the most comprehensive study of the northern spotted owl, Murphy and Noon
(1992) formulated a number of important, testable hypotheses regarding the
owl. All hypotheses listed below are null models.
1) Is the owl population growing (is lambda [finite
rate of growth] >1? Answer: Rejected. In both populations
studied (later confirmed with other populations), lambda was significantly
lower than 1 (replacement rate). Thus, the population is declining.
2) Owls do not differentiate among forests of
different ages or structures. Answer:
Rejected. The owls prefer habitats based with
old-growth forest in disproportion to the abundance of this habitat type
in nature.
3) Habitat type selected by the owls has not changed
in aerial abundance. Answer:
Rejected. See figure above. 70% of owl
habitat is currently on federal lands, and this habitat is
declining.
Based on these results, they wanted to devise
a protection scheme to stabilize the population by devising a Habitat Conservation
Area (HCA) that would, at a minimum, permit the owl to persist for at least 100
years. They thus constructed another set of testable hypotheses:
4) The probability of persistence is not related to
the extent of its geographic distribution. Answer: Rejected.
Spreading the risk of environmental catastrophes or disease outbreaks by having an extensive distribution reduced the chance
of species extinctions. The idea here is that extinction is less
likely to occur to species that occupy a large portion of their historic
range than a narrow portion.
5) No relationship exists between HCA size and its
owl carrying capacity. Answer: Rejected. Owl abundance was positively correlated with forest size, though forests of equal
size in different regions supported differing numbers of owls.
6) No relationship exists between HCA size (or carrying capacity) and population stability (likelihood
of population persistence). Answer: For British Island birds, persistence
time is positively correlated with island size.
7) No relationship exists between habitat
fragmentation and persistence likelihood of species using that habitat
landscape. Answer: Rejected. Fragmentation results in large edge
effects and reduced carrying capacity. Intact forest best promoted
growth of the spotted owl population.
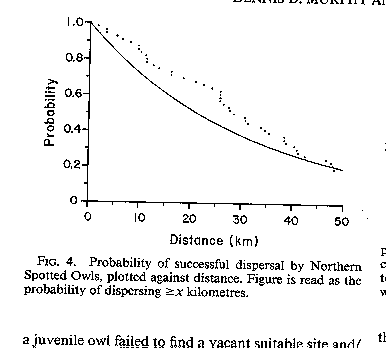
8) Distance between habitat patches has no bearing on
dispersal success of juvenile owls (i.e. no rescue effect). Answer:
Rejected. There is a very strong relationship.
See
Owl Dispersal (Murphy and Noon 1992)
9) Distance between HCAs
has no bearing on persistence likelihood. Answer: Rejected. Closer
spaced HCAs will foster metapopulation exchanges and
thus enhance survival likelihood.
10) No relationship exists between HCA size or shape
and carrying capacity. Answer: Rejected. A pronounced negative edge
effect was evident.
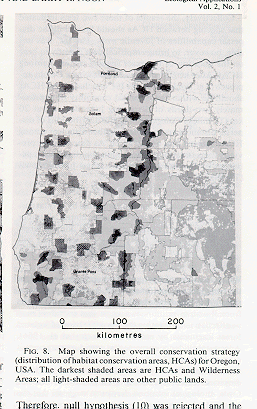
Based on this exercise
of hypothesis testing and available public lands which could serve as HCAs for spotted owls, the Spotted Owl working group
devised a patchwork of forests in the western USA to attempt to save the owl
(for 100 years).
See
Map of Oregon HCAs (Murphy and Noon 1992)
References
Bart, J. 1995. Amount of suitable
habitat and viability of northern spotted owls. Conservation Biology 9:943-946.
Bart, J. and E.D. Forsman.
1992. Dependence of northern spotted owls (Strix occidentalis caurina),
on old-growth forests in the western USA. Biological
Conservation 62:95-100.
Crooks, K.R., M.A. Sanjayan, D.F. Doaks. 1998. New insights on cheetah conservation
through demographic modeling. Conservation
Biology 12: 889-895.
Dunbar, D.L. et al. 1991. Status of
Spotted Owl, Strix occidentalis,
and Barred Owl, Strix varia, in
southwestern British Columbia. Canadian
Field Naturalist 105:464-468.
Laurenson, M.K., N. Wielebnowski
and T.M. Caro. 1995. Extrinsic factors and juvenile mortality in cheetahs. Conservation Biology 9:1329-1331.
May, R.M. 1995. The cheetah controversy. Nature 374:309-310.
Meffe, G.K. and C.R. Carroll. 1997. Principles of Conservation Biology. Sinauer, Sunderland, MA.
Menotti-Raymond, M. and S.J.
O'Brien. 1993. Dating of the
genetic bottleneck of the African cheetah. Proceedings of the National Academy of Science 90:3172-3176.
Merola, M. 1994. A reassessment of homozygosity
and the case for inbreeding depression in the cheetah, Acinonyx jubatus: implications for conservation. Conservation Biology 8:961-971.
Murphy, D.D. and B.R. Noon. 1992. Integrating scientific methods with habitat
conservation planning: reserve design for northern spotted owls. Ecological Applications 2:3-17.
O'Brien, S.J., D. Wildt, D. Goldman, C. Merril and
M. Bush. 1983. The cheetah is depauperate in genetic variation. Science 221:469-462.
O'Brien, S.J. and 9 others. 1985. Genetic basis for species vulnerability in the
cheetah. Science 227:1428-1434.
O'Brien, S.J. and 7 others. 1987. East African cheetahs: evidence for two
population bottlenecks? Proceedings of
the National Academy of Science 84:508-511.
O'Brien, S.J. 1994. The cheetah's
conservation controversy. Conservation
Biology 8:1153-1155.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}