Introduction to Biogeography
and Conservation Biology
The first requirement in any
subject is to define the limits and the connections in it. The textbook
standard definition of biogeography is that it is the study of the
distributions of species over space and time and the causes of those
distributions. It has two basic components: descriptive biogeography and
ecological biogeography. The first part studies the geography of species and
the second part deals in the causes of those distributions. Clearly the first
part is a necessary component, but it is the second part that is more exciting.
The subject as a whole is dynamic because its subjects - biological populations
and species- respond dynamically to causative factors over time and space, and
because it is a relatively young science.
The time scales which you will
see are important in biogeography range from days, weeks, or a few months, i.e.
periods we can describe as ecological time, to millions of years, which
constitute evolutionary or geological time scales. Physical events or changes
driving these dynamics range from rapid effects of nonindigenous species (also
called alien species) introductions, which may occur over short periods, to
continental drift and plate tectonics, in which the most recent continental
movements have occurred over the last 200-230 million years.
To indicate the dynamic
character of biogeography, we need only consider the recent history of the
subject, and the impact of the theory of continental drift on biogeography. The
concept of a plastic mantle for the earth, plate tectonics, and continents
moving, carried on the plates was only developed during the 20th
century. The theory was initially proposed by Wegener
in 1912. As recently as 1957 there is clear evidence that theory had not been
widely assimilated and accepted. One of the leading biogeographers of the 20th
century, Philip Darlington rejected the concept of continental drift in the first
version (1957) of his classic book.
Other biogeographers, as well as Darlington in a revised edition (1965),
rewrote ideas about explanations to include the impact of continental drift.
Today, no biogeographer questions the impact of drift.
Similarly, we can look back to
Darwin to appreciate his recognition of islands as important natural
experiments in ecology and evolution. However, island biogeography was
initially viewed as an interesting sidelight to larger problems of continental
distribution. Now island processes are seen as important in understanding the
population dynamics and genetics of species having patchy distributions, which,
at one scale or another, characterizes the distributions of almost all species.
Islands are now seen to include situations as diverse as oceanic or fresh water
islands or lakes in a terrestrial matrix, habitat islands, peninsulas,
woodlots, clumps of zebra mussels on lakebeds of the Great Lakes, and even
patches in the abundance distribution of species scattered widely over an area.
Lake Huron is an aquatic island on the North American continent, Manitoulin Island is an island in Lake Huron, and lakes are
biological islands on Manitoulin Island. Island concepts are also important in
considering how best to preserve species and communities as humans impose
themselves on natural environments. We
can trace the recognition of the broad importance of islands to important
publications by Robert MacArthur (1963) and Edward Wilson (in 1967).
The theory of island biogeography
is a critical component in the design of natural preserves and in assessing the
likelihood (or time) of persistence for endangered populations. However, even
as it is indicated as important, it should also be recognized that aspects of
the theory are still very contentious. There was an extensive literature,
mostly from the early 1980's, arguing whether the statistical tests of
observations based on the model, particularly occupancy and coexistence on
islands, were correct. Dan Simberloff argued that appropriate null hypotheses,
against which to test observations and model predictions, had not been
formulated. Related arguments about whether communities are random assemblages
from pools of potential colonists, or whether there are discernable assembly
rules that can predict which species (or at least species from particular
ecological groups) will be represented together in established
communities. Some aspects of
biogeography remain highly relevant to conservation programs today, although
the same issues can be addressed through studies of sub-divided populations,
called metapopulations.
The science of biogeography can
only realistically have begun when its students got past simply describing and
naming new species, and into describing habitat characteristics and
relationships among species (e.g. competition, predation, mutualism),
dispersal capabilities in relation to geological events, climate and climate
change, and continental drift. That is not an exhaustive list, but fairly
inclusive. The first attempts at this sort of analysis are traceable to
phytogeography in the first two decades of the 19th century. In 1805 Alexander
von Humboldt developed quantitative indicators of relationships between plant
species and climate, indicating an initial subdivision of climate types.
Humboldt made extensive collections of plants and associated environmental
variables throughout Latin America during a 5 year research trip, from tropical
forests to Peruvian alpine communities. That first classification was rapidly
developed further, particularly by DeCandolle in
1813, but summarized more thoroughly in Candolle
(1855).
Not long afterward, first Lyell,
then Darwin and Wallace, turned the world of geology, biology and biogeography
on its collective ear. Lyell proposed gradual change in the geographical
features of the earth. In the process, he developed the concept of uniformatarianism, the idea that
processes today are identical to processes operating in the past. It is that concept
that allows us to infer history from observations made in contemporary time. If
the forces were different, then science, and certainly biogeography, would make
little progress. Uniformatarianism does not state that the way things are
happening now is identical to the way they happened in the past, but rather
that the forces or processes that determine pattern today are identical to the
processes which drove changes in the past. Darwin and Wallace, in their
separate collecting trips in the new world and southeast
Asia respectively, observed species distributions which were critical to the
development of the theory of evolution. Darwin noted the presence of shells of
marine gastropods high in the Andes in southern Argentina and Chile, and
recognized that these areas must once have been marine
for shells to be found there. That is, a significant proportion of his
information was biogeographical. One of the results of the revision in
biological thought caused by the theory of evolution was a parallel revision in
biogeography, which could be argued to begin with the works of Ernst Haeckel. He named a special discipline called chorology,
which was the study of the spatial distributions of organisms and their causes.
One of the major components explaining the change in species' distributions was
evolution, and we now call the subject biogeography, which he called chorology.
We all know that one of the
driving forces which caused Darwin to publish his theory was the parallel
development of a nearly identical theory by Alfred Russell Wallace. Darwin is
regarded as the father of the theory of evolution, but both men made very
substantial contributions. Wallace and Darwin differed in their beliefs
concerning the forces which drove evolutionary change. We know Darwin's belief
about biological interactions driving evolution. Wallace felt that abiotic
forces were of great importance. One probable reason is differences in the
organisms whose evolution was studied. Wallace made his observations mostly on
insects, which are more likely to be affected by climatic change or difference.
Both Darwin and Wallace made numerous further contributions, but Wallace is
regarded as the father of zoogeography. Because animals are mobile, and because
so long was spent simply categorizing insects (because there are so many),
'animal biogeography' took longer to get off the ground. The earliest, still
valuable animal biogeography is Wallace's master work, The Geographical
Distribution of Animals. In it Wallace described the transition in terrestrial
fauna between Australia, with its associated islands, and the islands extending
out from southeast Asia towards Australia. The
exchange of fauna (excepting animals capable of flight
or transferred by man) is extremely limited. Two lines have since been drawn by
later biogeographers to demark this transition. Weber's line encloses the
region in which the mammalian fauna is exclusively Australasian, and west of
the Celebes, between Bali and Lompok, is Wallace's
line, which marks the outer limit of the Asian (or Oriental) mammalian fauna.
In the narrow zone between these boundaries there is limited mixing; the area
is called Wallacea.
Figure
1 - A map of Austalasia, with the positions of Weber's
and Wallace's lines marked.
It could be argued that the 60-70 years following Wallace's
zoogeographic work represented a period of consolidation, of data gathering,
which was the basis for the radical developments of the last 20-30 years. That,
however, would miss various important contributions which occurred in the late
19th and earlier 20th centuries. Among these are included:
1. Bergmann's rule which states that warm blooded animals from
cooler climates have larger body sizes
and lower surface to volume ratios. That change in body plan is clearly
adaptive to restrict heat loss in cooler climes, and to maximize heat
dissipation in warmer areas. A good comparison to indicate the difference is to
compare the body form of arctic hares (larger bodies, generally 'more
spherical') with those of temperate cottontails or jackrabbits (smaller bodies,
on average 'longer and leaner').
2. Allen's rule which says that warm blooded animals will have
more compact extremities in cold climates than warm ones. The underlying
reasoning is the same, i.e. optimization of design for heat dissipation in
relation to climate. Following the same comparisons, limbs and ears are shorter
in the arctic hare, longer and thinner in the cottontail, and notably long in
the jackrabbit.
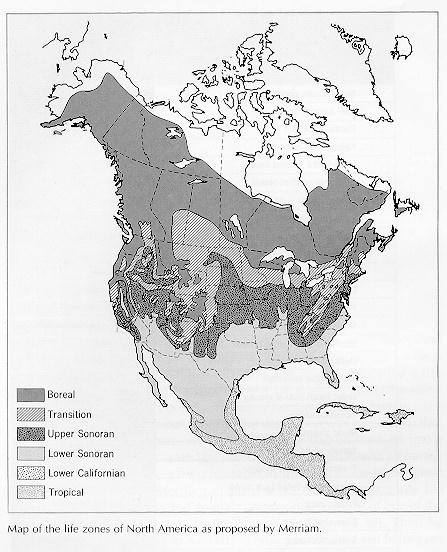
3. Merriam's classification of altitudinal and latitudinal
vegetation types and zones, termed life zones, and their relationship to
temperature and rainfall. Most modern diagrams of Merriam's zonation
present it as a three axis system, in which the climatic axis is potential
evapotranspiration. This represented an advance on the earlier plant
biogeography. Merriam attempted to generalize his classification scheme to
animals, and failed.
Figure
2 - Merriam's life zones.
Finally, we come to the
quantitative theory of biogeography, traceable to the monograph by MacArthur
and Wilson. While controversies have since developed about the breadth with
which the basic theory can be applied, much of its basic structure deserves the
same comment that Huxley made upon reading Darwin's The Origin of Species:
"stupid of me not to think of that myself".
The history of conservation
biology parallels that of biogeography to at least the degree that they
overlap. There are unique aspects, however. In this case we are interested in
the history of extinctions and the recognition of human impact. Humans (native Americans and Inuit) are widely regarded as
responsible for the extinction of a variety of large mammals in North America
including mastodons, tapirs, glyptodonts, and giant ground sloths. Humans have
long cut forests. In Greek times, the forests of the Baltic area and those in
southern Asia were cut for ship building. Tropical forest has been cut for
centuries in the course of slash and burn agriculture. When one patch used for
subsistence agriculture gave out, a farmer moved to a nearby patch, cut the
trees down, then burned the logs, releasing the nutrients tied up in the
biomass. Until the soil hardened (laterization) and nutrients percolated down
or washed away, the farmer used the patch. When yield dropped, he moved on
again. The contemporary problem is the effect of increased population size,
meaning larger areas cut and more frequent exhaustion of areas, combined with
other impacts mentioned earlier.
There are some common themes.
One of the most important is the 'tragedy of the commons'. If everyone takes
advantage of common, public areas, each thinking his or her small impact is not
important, the summed result is the destruction of the
commons. Garrett Hardin, in one of the seminal papers of conservation ethics,
wrote about the occurrence of exactly this phenomenon in the commons of New
England towns, where grazing one extra cow was thought to be insignificant. The
same thing happened in Europe, where royal preserves and the manor lands of the
wealthy were unavailable, and the public lands (the commons) were deforested to
provide charcoal for heating and industry. There the industrial revolution was
the last straw, and Great Britain was largely deforested by the end of the 18th
century. It is not that the conservation
ethic has not been recognized for far longer than the discipline has existed;
it is that scientific efforts to develop a framework for broad principles of
conservation have only developed recently.
Conservation biology is a much newer discipline than biogeography. It is
always difficult to set a time of origin for something as abstract as a
discipline, but the best guess would be to say that conservation biology came
into existence as a distinct discipline with the recognition in the 1970's that
the rate of extinction of species globally, partially to largely due to the
influence of human activities and population growth, is now high enough to
parallel rates which were previously only 'seen' during some of the megafaunal
extinctions of the past. Therefore, conservation biologists describe the
current scenario as a "biodiversity crisis". The rates they quote are
almost certainly conservative and lower than actual rates of species loss. The
reason is that the areas of greatest diversity, e.g. tropical rain forest and
coral reefs, are also areas with the greatest numbers of as yet undescribed
species and simultaneously areas under severe pressure from human activities.
Tropical rain forest is being cut for fuel wood, lumber, and conversion to
pasture land at a rate which will lead to the elimination of all but small
protected reserves within about the next 50 years. Coral reefs are affected by
physical and chemical degradation due to human activities on nearby terrestrial
areas.
Conservation biology is a
synthetic science, built from ecology, population biology, population genetics,
biogeography, economics, anthropology, philosophy, and probably other
disciplines with the intention of developing principles and strategies to
preserve diversity. Different approaches may attempt to maintain the diversity
of species directly, or through maintenance of a diversity of habitats. It must
maintain a balance between the potential desire to preserve everything in a
pristine natural state, and the political desire to permit intensive development.
The buzzwords among those who work hardest at rational balance are
"sustainable development", though there is not yet an established
theoretical basis for it.
The main objectives of Conservation Biology are to:
·
Human population growth has accelerated during the
past 400 years, and 100 years in particular; and resource demands have risen
accordingly;
·
Present threats to biodiversity are unprecedented;
never in human history have so many species and habitats been threatened;
·
Many of the threats to biodiversity are synergistic
(additively or multiplicatively)
o
e.g. eutrophication (i.e.
nutrient enrichment) and overfishing;
o
e.g. species invasion and climate change (e.g. malaria
in North America)
o
species invasion and overharvesting: e.g. extinction on Feb. 1, 1996 of
Polynesian tree snail (Partula partula).
This species succumbed to combined pressures of human exploitation of its
pretty shells, and a nonindigenous mollusc predator introduced for biocontrol of a different species;.
o
Environmental deterioration may signal pending human
misery
1) e.g. history of humans and environmental
disaster that occurred on Easter Island (south Pacific)
2) recent history of the Aral Sea. Salinity of this inland
'sea' has increased tremendously as humans appropriated (diverted) water
inflows for agriculture irrigation purposes.
As the basin's surface area and volume have declined, it has become
increasingly inhospitable to human usage (e.g. as a source of fish).
Conservation Biology is a very
new discipline; its most prominent journal
Conservation Biology was created in only 1987 and the Society for Conservation
Biology (created by Michael Soulé, Paul Ehrlich and Jared Diamond) was founded
in 1985. The society has grown explosively since then.
Basic Principles of Conservation Biology (Meffe and Carroll 1997)
These principles are largely an
offshoot of the statement by the famous ecologist G.E. Hutchinson in which he
stressed the 'ecological theater and the evolutionary play'. Ecological
events help shape evolutionary patterns of species and communities; in order
that evolution be permitted to occur more or less unencumbered, we must not
destroy the habitats and species that facilitate evolution.
Richard Primack (1994) has also established basic principles:
Candolle,
A. de. 1855. Géographie botanique
raisonée. 2 vol. Paris, Masson Editeur.
Clausen, Keck, and Heisey. 1948.
Experimental studies on the nature of species. III. Environmental responses of climatic races
of Achillea. Carnagie
Institute, Pub. no.581.
Darlington, P. 1957. Biogeography of the Southern End
of the World. Harvard Univ. Press, Cambridge, MA.
Hardin, G. 1968. The tragedy of the commons. Science
162:1243-1248.
MacArthur, R.H. and E.O. Wilson. 1963. An
equilibrium theory of insular zoogeography. Evolution 17:373-387.
MacArthur, R.H. and E.O. Wilson. 1967. The Theory
of Island Biogeography. Princeton Univ. Press. 203p., Princeton, NJ.
Meffe, G. and C.R. Carroll. 1997. Principles
of Conservation Biology. Sinauer, Sunderland, MA.
Primack, R.B. 1994. Essentials of Conservation
Biology. Sinauer, Sunderland, MA.
{kind=link}
{kind=link}